Biological Design for Lungs and Gills: Biomechanics and Materials
Samuel Bernard, Nabhaan Farooqi, Camille Gagnon, Will Vyse
Abstract
This report describes two of the most important gas exchangers for all living animals: lungs and gills. Throughout the research, cutaneous respiration will also be explained; however, the most efficient oxygen diffusing rates are obtained through lungs and gills, which are our main concern. The purpose of this biological design project is to find out how different species evolved and developed unique respiration systems that allow them to maximize their oxygen intake without spending energy. This report contains equations derived from fluid mechanics to explain viscosity resistance, the effect of surface area for both gills and lungs, and the correlation between the surface area and the animal's body mass.
Evolution of Oxygen-consuming Organisms
Terrestrial and aquatic animals both breathe in their respective environments, due to a very simple process: gas exchange. In the earliest period of life on Earth, anaerobic unicellular organisms lived in an anoxic environment. Through redox pathways, these cells were able to find enough energy to power their cellular process without oxygen. These “redox reactions” release energy as electrons transfer from a molecule to another, thus meaning that these reactions require main electron donors to fulfill the electron donor-acceptor cycles: such as H2, H2S, CH4, and nitrogenous compounds. Water was also present on Earth; however, using water as an electron donor was extremely difficult since none of the microorganisms had the energy to split the ions. For a billion years, these cells had to rely on the redox pathways to survive before a specific bacteria managed to evolve into stronger cellular organisms able to divide water and use it as an electron donor for photosynthesis. To mammals, oxygen is the most essential gas in the atmosphere as it allows them to breathe. However, at that time, it was a powerful toxin to most organisms. The effect of oxygen on the microorganisms was devastating since it affected their redox reactions. To survive, specific cells, aerobic prokaryotes, were able to consume oxygen as a new source of energy. Research and experimentation have proven that aerobic respiration provides organisms with greater amounts of energy than anaerobic respiration. Bacteria that took advantage of this and utilized the oxygen typically could produce sixteen times more energy than those which could not. As a result, the aerobes multiplied faster than the anaerobic cells. Some of the larger bacteria, usually anaerobes, would engulf smaller prokaryotes and use them as gas-consuming compartments. These smaller prokaryotes would then evolve into mitochondria, small organelles which manage to use oxygen in a process called cellular respiration, whereby glucose, a sugar, is used to synthesize ATP, an energy rich form of fuel for the cells. These larger organisms are known to have become eukaryotic cells, of which all animals are composed.
Further along the timeline, variations between oxygen tension and various environments, water, and earth, allowed microscopic organisms to evolve and develop unique respiratory systems using different gas exchange mechanisms: gills, lungs, and for some aquatic species, skin. At that time, cutaneous respiration was the most abundant gas exchanger, as it offered a larger surface for gas diffusion. However, today, many organisms have developed to be much larger and for some, the diffusion rate of oxygen through the skin compared to the typical rate of oxygen consumption would be simply too slow for cutaneous respiration to be the only reliable method of gas exchange. Nature had to evolve and develop respiratory systems that could adapt to their size, such as lungs and air sacs. Furthermore, depending on their environment, specific organisms can even possess temporary gas exchangers. For example, most amphibians, as larvae, use two different gas exchangers, gills and skin, but are usually born with non-operational small air sacs that serve as lungs. However, during the metamorphosis climax, these lungs start to develop, and their gills degenerate. There are, nonetheless, some exceptions to the rule, such as a unique class of salamander, which possessing no lungs or gills, may breathe through only the pores on its skin.
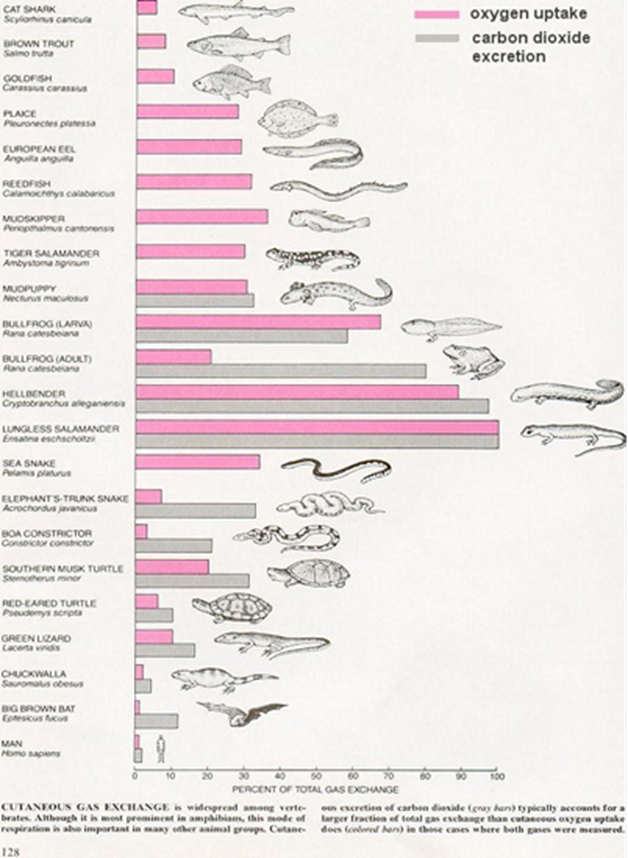
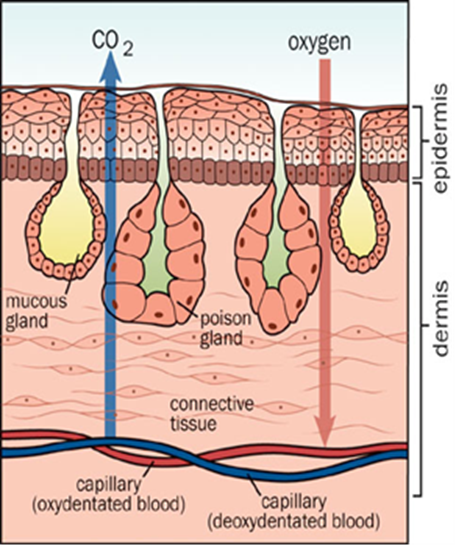

Gas Exchange: A General Overview
By definition, the kinetic energy of gas molecules is much greater than their intermolecular forces of attraction which allows them to move in random directions inside a container. However, gas particles will follow the same direction and always diffuse toward an area that possesses a lower concentration of gas. The rate at which the gas diffuses can and will be affected by a few factors, such as the gradient of concentration, the surface area for diffusion, and the length of the diffusion pathway. Although we will explain this subject in greater detail further along, essentially, if there is a greater difference in the concentration of gas between two areas, then the diffusion rate will be higher. Additionally, if the surface area for diffusion is larger, then the diffusion rate will also be higher. If the diffusion rate increases, the number of oxygen molecules that can access the bloodstream follows. Gas exchangers all have similar components: a thin blood-gas barrier, a large interface, and ventilatory regulation. This gas exchanging process occurs in gills, lungs and skin.
Gas Exchange in the Lungs
Gas exchange occurs at two sites in the mammal body: at the lungs where oxygen enters the body and carbon dioxide is released and at the blood capillaries of tissues where oxygen is released and carbon dioxide is released, as a result of cellular respiration. The respiration that happens at the lungs, more precisely between the alveoli and the blood in the pulmonary capillaries is a function of partial pressure differences in oxygen and carbon dioxide. Both gases move independently of each other and diffuse down their own pressure gradients because the rate of diffusion of a gas is proportional to its partial pressure within the total gas mixture. Gas transport therefore relies on a passive diffusion and is influenced by some determining factors.
Fick's Law
According to Fick's law of diffusion through fluid, diffusion of a gas across the alveolar membrane first increases with the surface area of the membrane. Since oxygen has a relatively low solubility, a large internal surface area is necessary for the tissue cells to obtain all the oxygen they need. Thus, many alveoli are present in the lungs with a shape that further increases surface area. The spherical or grape-like shape of the alveoli allows for maximum contact between the alveoli and the capillaries that surround them. Because of their structure, the alveoli of a human are spread on a surface area of approximately 75 m2. Another factor influencing the gases' rate of diffusion is the length of the diffusion pathway. The thinner the respiratory surface is, the faster the diffusion will occur. To facilitate diffusion, both alveoli and pulmonary capillaries are only made of one layer of cells. Combined to the other diffusion barriers, this means that the air in the external environment is only separated from the internal environment, meaning the blood, by the alveolar epithelium, the tissue fluid and the capillary epithelium. For oxygen, once it is dissolved in the blood, it also has to go through plasma and red cell membranes, where it will be fixed to the protein hemoglobin, which provides transport of oxygen throughout the capillary system.
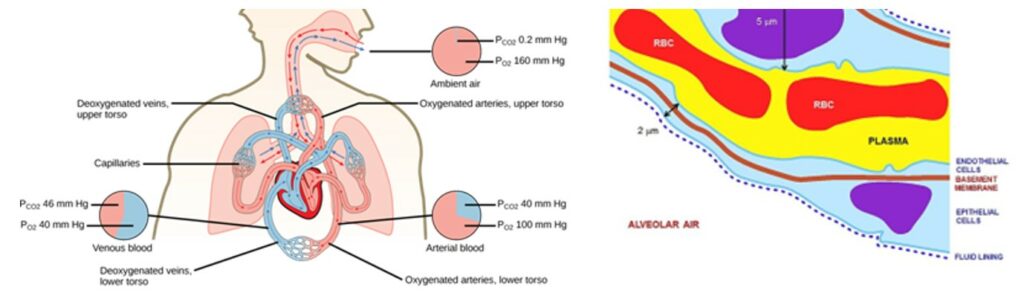
Besides membrane thickness and surface area, Fick's law of diffusion also mentions that pressure difference across the membrane and diffusion coefficient of the gas, a property specific to each gas, have an impact on the rate of diffusion. To summarize, considering the different factors, Fick's law can be expressed mathematically as:
V_{gas} = (A \times D \times (P_1-P_2))/Twhere:
- Vgas = volume of gas diffusing through the tissue barrier per unit time (mL/min)
- A = surface area available for diffusion (70 m2 in a healthy adult)
- D = diffusion coefficient, or diffusivity, of the particular gas. For example, CO2 diffuses approximately 20 times (24/1.17) more rapidly through the alveolar-capillary barrier than O2.
- P1 – P2 = partial pressure difference of the gas across the barrier
- T = thickness of the barrier or the diffusion distance (0.2 to 0.5 mm)
Lung Capacity
To complete gas exchanges, lungs do not usually have to fill to maximal capacity. According to Jonathan P. Parsons, M.D., professor of internal medicine, associate director of Clinical Services and director of the Division of Pulmonary, Allergy, Critical Care and Sleep Medicine at the Ohio State University Asthma Center, “The lungs are over-engineered to accomplish the job that we ask them to do. In healthy people without chronic lung disease, even at maximum exercise intensity, we only use 70 percent of the possible lung capacity” (Wilkinson, 2017). Lung capacity is important and predicts health and longevity. Athletes, for example, usually have a greater lung volume because their body must be able to extract more oxygen. Consequently, lung capacity can be improved but lung function, which refers to how effectively your lungs both oxygenated and remove carbon dioxide from your blood, cannot. Human lung size is determined by genetics, gender, and height. At maximal capacity, an average lung can hold almost six liters of air but this number decreases with age. Among other mammals, the lung capacities are also different based on the activities they execute. Cheetahs, for example, have evolved a much greater lung capacity than humans to allow them to run very fast by providing oxygen to all the muscles in the body.
Pleural Membranes
Nevertheless, the primary role performed by the lungs is gas exchange because the organs themselves are passive during breathing since the movements of the lungs' membranes relies on muscles of the thoracic walls. Indeed, the outside of the lungs is formed by the pleural membranes, which consist of serous membranes that fold back onto themselves to form a two-layered membranous structure. The two layers are the outer pleura (parietal pleura) attached to the chest wall and the inner pleura (visceral pleura) covering the lungs and adjoining structures. The thin space between the two pleural layers is the pleural cavity and normally contains a small amount of pleural fluid. The parietal pleura is highly sensitive to pain, while the visceral pleura is not, due to its lack of sensory innervation. Pulmonary ventilation is completed in two steps: the inspiration and expiration where the pressure gradients are created and drive the air outside or inside the lungs.
Intrapleural Fluid Force
During inspiration, the expansion of the chest wall causes both lungs to stretch out due to the parietal pleura being attached to the ribs and diaphragm and to the adhesive force of the pleural fluid. This force occasioned by the intrapleural fluid that holds the two membranes together is crucial because there is no anatomical connection such as blood vessels or connective tissue between the two pleural membranes. In fact, they are not supplied in blood by the same circulatory system of blood vessels. The adhesive force can be compared to two microscope slides with droplets of water between them. It is easy to slide them horizontally over one another but pulling them apart perpendicularly is very difficult. The same concept applies when the chest expands: the lungs slides against the thoracic wall and lung volume increases. In other words, the outer membranes of the lungs play a very important role in the gas exchanges but the overall structure of the lungs is much more complex.
Conducting Zone
Before diffusing through the membranes and arriving in the respiratory zone, air follows a pathway throughout the different structures of the conducting zone. Following the order from the external environment to the internal environment, the nostrils, pharynx, larynx, trachea, bronchi and bronchioles have hair, cilia and mucus that warm up, humidify and purify air. Making its way through the trachea and entering the lungs, air is led through the bronchial tree, the very complexed branching structure of the bronchi, which represents the conducting zone of the organ. It develops by branching of the epithelium into the surrounding mesenchyme, a source of specification that provides critical signals for respiratory lineage specification.
Branching Process
The bronchial tree of a human has more than 105 conducting and 107 respiratory airways arrayed. The knowledge regarding the program that directs branching configuration is still rudimentary. However, according to a study conducted by searchers from Department of Biochemistry of Stanford University and Department of Anatomy or the University of California (Metzger et al., 2008) on reconstructed mouse lungs, the pattern of branchial three is stereotyped in three branching methods presented on Fig. 3 that exist repeatedly in the lungs: domain branching, planar bifurcation and orthogonal bifurcation.
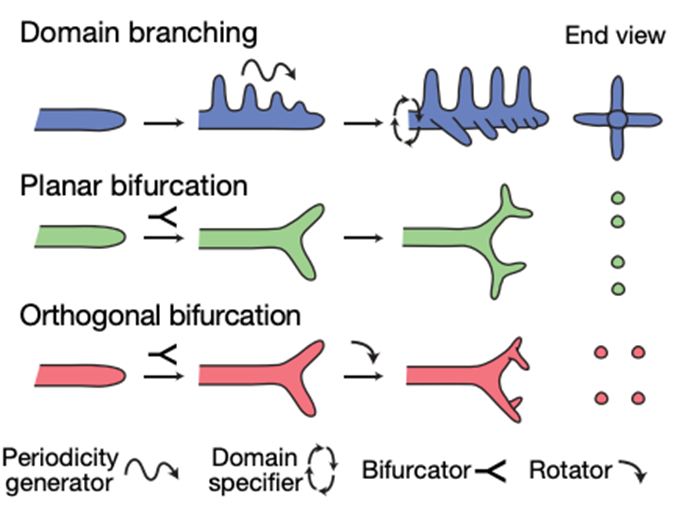
The first one, domain branching is controlled by two patterning systems, known as a proximal-distal system that controls the sequence of each branch on the parent branch and a circumferential system to specify the position and order in which domains (each tiny branch) are used. As for the second and third methods, planar and orthogonal bifurcation mostly form the tertiary and later-generation branches. The mode of branching towards the end of the bronchial tree is rather a procedure where the tip expands and bifurcates. For planar bifurcation, all tip divisions occur in the same plane whereas for orthogonal bifurcation, between each round of branching there is a 90º rotation in the bifurcation plane. This results in four granddaughters arranged in a circular formation. During the first five days of lung development, the three branching modes are used at several different positions and each mode is associated with a specific aspect of lung design. As the result of the study conducted by searchers from Department of Biochemistry of Stanford University and Department of Anatomy or the University of California: “Domain branching is used first and generates the central scaffold of each lobe, setting its overall shape (for example, trigonal pyramidal). Planar bifurcation forms the thin edges of lobes, and orthogonal bifurcation creates lobe surfaces and fills the interior” (Metzger et al., 2008). Even if there is some variance in the branching process, the branching program behind the resulting pattern is very logical. It thus proves that lungs are a well-evolved and well-engineered system.
Description of Gills
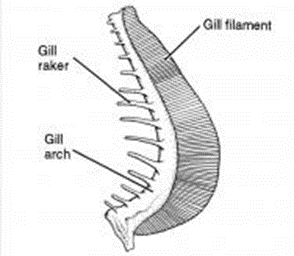
Gills work by bringing oxygen into fish's blood, and removing carbon dioxide. They do this by interacting with water. Water enters through a fish's mouth, goes to the gills, exchanges oxygen and carbon dioxide with the fish's blood, then returns to the environment, leaving through the gills. Some fish need a continuous flow of water to oxygen. These are the fish that are constantly swimming, like tuna. Other fish can pass water over their gills by pumping it. These are the fish that generally remain stationary, such as wrasses. They are composed of three organs, the gill filaments, the gill arches, and the gill rakers.
Primary and Secondary Lamellae
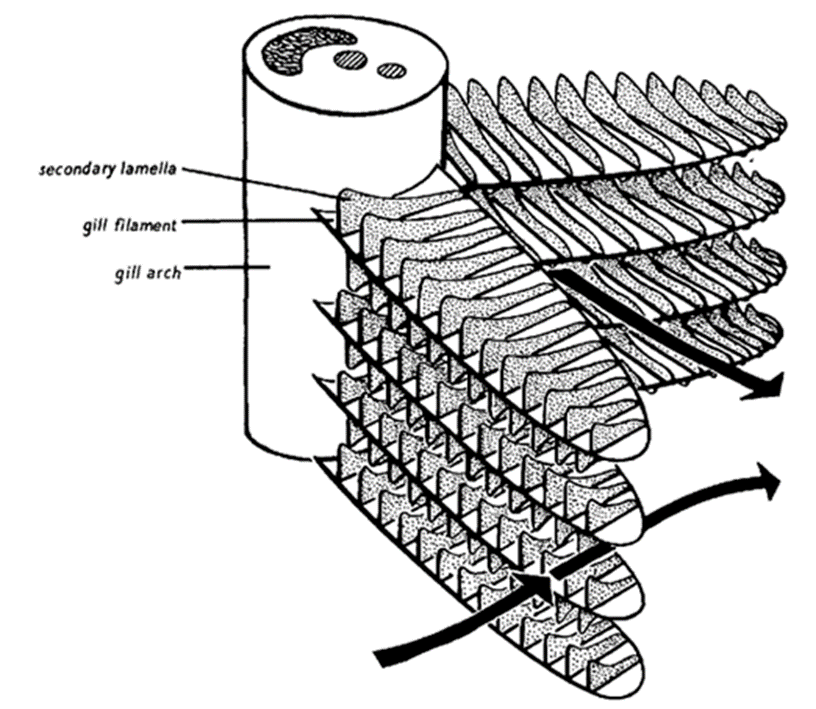
The gill filaments, also known as the primary lamellae, are intricate structures with large surface areas. They are soft, and are primarily made up of blood vessels. They also contain small offshoots, called secondary lamellae, which contain blood capillaries and are the ones responsible for absorbing oxygen into the blood, and diffusing carbon dioxide from the blood to the water. Fish that are active and frequently swimming have highly developed gill filaments, to maximize the absorption of water, while passive fish that live on the ocean floor usually have smaller gill filaments, because they are less active and do not use up oxygen as quickly.
Arches and Rakers
Gill arches are bony, boomerang-shaped organs. They consist of an upper and lower limb that are joined in the back, and are attached to both to the gill filaments and gill raker. Most fish have at least three gill arches on each side of the body. Gill arches offer support for the gill, sort of like the gill's spine, keeping it from collapsing in on itself.
Gill rakers allow the fish to get energy by helping them eat. Food enters the fish from the mouth, just like the water that must flow to the gills. However, the food must go to the stomach instead. Gill rakers are what enable fish to divert food away from the gills, by being small enough to block off the path from the mouth to the gills, while still allowing water to flow into the gills. The number and shape of the gills depends on the diet of the fish. Fish that eat smaller prey, such as plankton, have many gill rakers which are long, thin, and close together, to prevent smaller prey from entering (and escaping via) the gills. On the contrary, fish that eat large prey would have smaller amounts of widely spaced gill rakers, as they do not need to catch the small particles.
Biomechanics of the Gill Surface Area for Diffusion
As mentioned above, water contains a much smaller amount of dissolved oxygen compared to air. This means that, to maximize the oxygen that diffuses into the bloodstream, the surface area (SA) for diffusion must be big enough. This SA is called the Gill surface area. Mathematically, it can be determined as a product of multiple factors:
GSA = TFL \times 2Nlam \times Alam
where GSA is the gill surface area; TFL is the total length of all the gill filaments, we calculate it by multiplying the average filament length by the number of gill filaments; Nlam stands for the number of lamellae per mm on a single side of the filament. Given that there is also a secondary lamella, Nlam is multiplied by a factor of two; Alam represents the average bilateral surface area of the secondary lamella. The product result is usually given in cm2. Consequently, if any of these factors increased, the GSA would also be bigger.
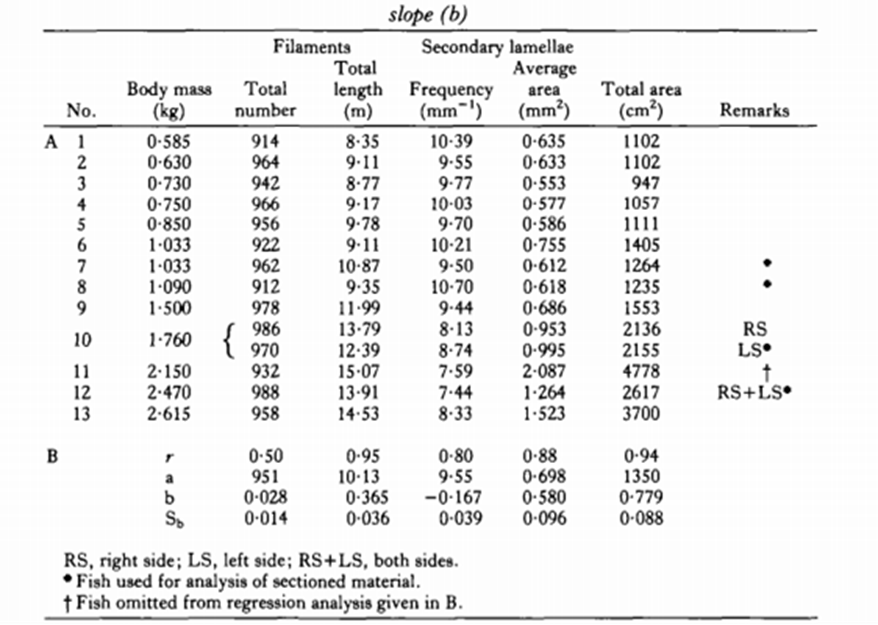
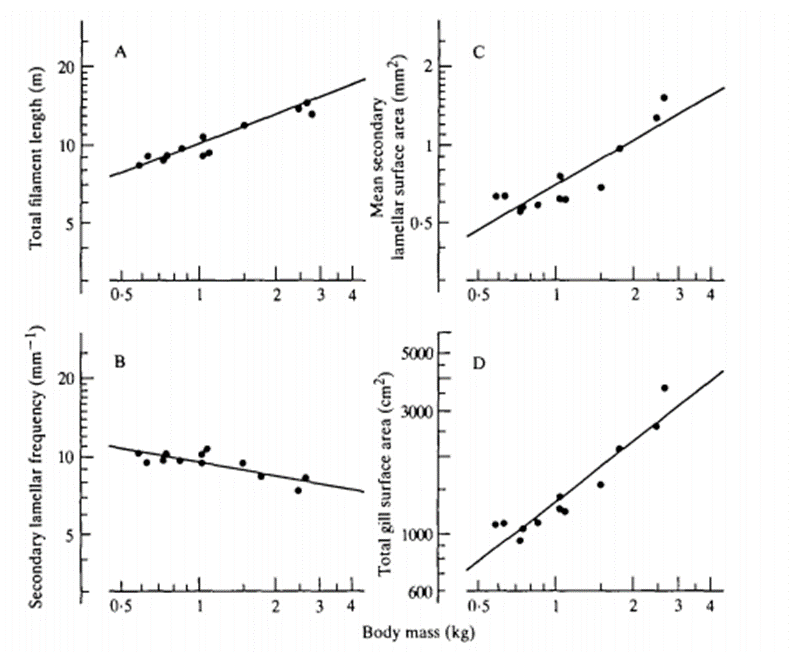
Correlation between the Gill Surface Area and Body Mass of Fish
After calculating the GSA, researchers discovered that there was a correlation between GSA and the fish body mass. As stated in the introduction, at their larval stage, some aquatic species will mostly take advantage of cutaneous respiration even if their gills are developing as it offers them a larger surface area for diffusion. However, as these organisms grow, the distance of diffusion between the bloodstream, the skin and the dissolved oxygen thickens which limits their gas exchanges. As the diffusion rate for cutaneous respiration decreases, their gills' diffusion capacity continuously increases. Once they mature, the distance of diffusion between their gills and the bloodstream slightly shrink and their gill surface area per body mass is at its highest. This is where the correlation is present, the total gill surface area is relative to the body mass, such that:
Log(Y) = Log(a) + (b) Log(W)
When looking for the total surface area, “a” is the equal to Y when W = 1 g, where W is the mass in grams and b is the allometric exponent, also referred to as the slope (see Fig. 7). If both sides of the equation are raised as exponents of base ten, the logarithmic components can be removed, simplifying the equation. The correlation between a and W is seen as:
Y=a \times W^b
The slope b is always between 0.36 to 1.13 for the gill surface area, which means that the Y value will continuously increase as the body mass increases.
Viscosity Resistance
Having a larger GSA is one way for aquatic animals to maximize their gas diffusion rates into the bloodstream, still there exist multiple other factors that allow some fish to diffuse oxygen easier than others. When water passes through the channels of the gills to contact the lamellae and let the dissolved oxygen enter the capillaries, these interlamellar channels create a viscous resistance due to the fact that the lamellar structures are held tightly together. This resistance affects the ventilatory-flow conditions in the channels applied by the fish. In fact, because of the way fish move, oxygen-rich water enters the gills continuously. Hence, the smaller the resistance is, the higher the gas diffusion rates will be. The viscous resistance is determined with the Hagen-Poiseuille equation, which explains the variations of water flow going through parallel plates:

R = 12 \mu l / d3h
where R is the resistance, µ signifies the dynamic viscosity of the water, l is the length of the interlamellar channels, d is the diameter of the channels and h is the height of the lamella.
The formula above only describes the resistance for a single interlamellar channel. For multiple channels, the equation would have to be modified, such that the variables w, the width of the lamella, and TFL, total length of the filaments, are added:
R = 12 \mu l (d+w) / d3hTSL
After reviewing this equation, it is clear that higher water viscosity, interlamellar channel length and lamellar frequency would increase the viscosity resistance limiting gas diffusion into the bloodstream and raise the amount of energy consumed in order to allow better ventilatory-flow conditions. On the other hand, an increase in the total length of the filaments and/or the height of the lamella would allow the fish to lose less energy trying to maximize their diffusion rates.
The Diversity of Life and the Role of Energy
Life on Earth exists in a truly extensive variety of forms: from unicellular prokaryotes, such as Pelagibacter ubique, which can be as small as 370 nm in length, to blue whales, which can span up to 30 meters. Despite the vast diversity of life, all lifeforms share specific common traits. One such trait is the fact that all organisms are composed of cells, the smallest structural unit of an organism that is still capable of independent functioning. Additionally, all cells require a source of energy to survive and perform all their necessary functions. Cells acquire this energy from the surrounding environment. Some organisms do this via a process called photosynthesis, whereby energy from the Sun is used to produce high energy sugar molecules from water and carbon dioxide. Other organisms, including all animals, acquire essential nutrients by consuming other organisms and metabolizing some of the cellular nutrients stored in them. Most cellular nutrients are relatively complex organic molecules, such as sugars, fats, and proteins, which contain large amounts of energy stored in their chemical bonds, making them essential as raw materials from which most lifeforms derive their energy. Ultimately though, the Sun is the original source of most cells' energy, since photosynthetic organisms such as plants and algae make up the base of most food webs.
Catabolic Oxidation Reactions: A General Overview
Once a cell has acquired high energy nutrient molecules, such as glucose, it cannot simply break these molecules and gain the energy. Cells release the energy stored in the nutrients through a series of oxidation reactions. Oxidation reactions are chemical reactions in which electrons are exchanged from one molecule to another. This results in changes to the composition and energy content of both the donor and acceptor chemicals. Nutrient molecules, such as glucose, are electron donors, which lose energy to electron acceptor molecules during each oxidation reaction. As a complex nutrient molecule is oxidized, it breaks down into simpler separate molecules, releasing energy, which is a process known as catabolism. If oxygen is present, the cells of eukaryotes and some prokaryotes can oxidize the carbon atoms in nutrients to the extent, whereby all of the carbons in the structure are separated and released as waste, in the form of CO2. As a result of oxidation reactions, a cell may use the energy once stored in nutrients to synthesize molecules such as adenosine triphosphate (ATP), which store high levels of energy in a readily-accessible format.
Aerobic Respiration, Anaerobic Respiration, and Fermentation
The catabolic four-step process of oxidizing nutrients in the presence of oxygen is called aerobic cellular respiration. In aerobic respiration, molecular oxygen (O2) is used as the final electron acceptor. Under conditions in which little to no oxygen is present, some organisms (usually prokaryotes) can use a different final electron acceptor, such as sulfate (SO42-) or nitrate (NO3–). This process is called anaerobic respiration. Since oxygen has a greater reduction potential than the alternatives, aerobic respiration releases more energy per oxidized molecule than anaerobic respiration. Lastly, in some cases, only glycolysis, the first of the four steps of respiration, occurs. This is known as fermentation, and it does not require the presence of oxygen. Fermentation usually involves the release of larger byproducts than respiration, due to the fact that far less oxidation occurs. For example, when the skeletal muscles in the human body do not receive enough oxygen during physical exertion, fermentation occurs, producing lactic acid, a three carbon byproduct, rather than CO2, a one carbon byproduct. Additionally, fermentation lacks the steps in respiration which result in a majority of the ATP synthesis, so it is an inefficient process, producing even less energy than anaerobic respiration.
Respiratory Mechanisms
Respiration in Unicellular Organisms
Organisms which perform aerobic and anaerobic respiration need to be able to take in adequate quantities of final electron acceptors (O2 in the case of aerobic respiration) and release the resulting waste products. In an unicellular organism, such as a paramecium (a eukaryotic protist genus) this process is very simple. Oxygen, the oxidizing agent and final electron acceptor, diffuses into the cell if the outside environment has a higher concentration of it. Then, as the concentration of CO2 increases, it diffuses out of the cell, into the surrounding environment where the concentration is lower. A paramecium uses aerobic respiration, but the process of in-taking the final electron acceptor and releasing waste products is fairly similar in organisms that use anaerobic respiration, too. Take for example the bacteria Paracoccus denitrificans, which can survive in anaerobic and aerobic environments. One of its many possible final electron acceptors is nitrate (NO3–), which is diffused into the cell. Nitrogen gas (N2) is the waste product of this process and it is diffused out of the cell. This type of anaerobic respiration is referred to as denitrification.
Multicellular Organisms – The Respiratory System
In multicellular organisms, a structural system may be required to effectively exchange oxygen and carbon dioxide with all of the cells. Such a system is known as a respiratory system. Respiratory systems vary greatly between species and the structures of which are dependent upon the habitat, size, and evolutionary history of the organism. A typical respiratory system requires a large, thin-walled, moist, and vascular surface to act as an interface for gas exchange to occur. In some organisms, this interface is referred to as the respiratory surface. The surface area of the interface is a major factor affecting the rate at which gas exchange can occur in an organism. The greater the surface area, the more oxygen an organism can intake and the more carbon dioxide it can expel. Larger or more active organisms require a greater rate of gas exchange and so require a larger respiratory surface. The respiratory surface must be quite thin, since this helps to increase its permeability to O2 and CO2, by reducing possible diffusion barriers. The thickness of a respiratory surface is referred to as the air-blood diffusion distance in terrestrial systems and the water-blood diffusion distance in aquatic systems. The purpose of moisture on the interface is to help dissolve gases in liquid, allowing them to pass into the circulatory system more easily. In most aquatic respiratory systems, this is already accomplished, due to the fact that the oxygen is already dissolved in the surrounding water. Lastly, the reason the respiratory surface is highly vascular is so that a large supply of oxygen can continuously be delivered to the body cells and carbon dioxide can be continually transported from the cells to the respiratory surface for release. Generally, the more vascular a respiratory surface is, the greater the amount of oxygen that can diffuse in and the amount of carbon dioxide that can diffuse out, at any given time. In plants, the respiratory system may work in two directions: intaking O2 for cellular respiration and expelling CO2, or in-taking CO2 for photosynthesis and expelling O2.
Cutaneous Respiration – Jellyfish
Cutaneous respiration is a method of gas exchange, whereby the respiratory surface is the outer layer or cuticle of the organism. In some multicellular organisms that do not require large amounts of oxygen, cutaneous respiration is a sufficient method of gas exchange. Organisms that rely only on cutaneous respiration tend to be simple in structure, have a low metabolism, dwell in aquatic environments, have a small water-blood diffusion distance, or have a high external surface area to volume ratio, typically meaning that they are quite small.
In the case of Malo kingi, a species of jellyfish, all of the characteristics that allow for cutaneous respiration are present. Firstly, as a jellyfish, it is rather simple in structure, possessing no conventional organ systems, which reduces the amount of oxygen that its cells need. Additionally, jellyfish like Malo kingi do not require a particularly high metabolism, since their behavior does not entail any energy-intensive activities, and their movement is generally quite slow. All cnidarians have bodies that are made up of only two cell layers which help to decrease the diffusion distance between water and the body cells. Jellyfish have no circulatory system to transport oxygen around the body, since the folded shape of the jellyfish allows for a large enough surface area to volume ratio such that all of the body cells are close enough to the external environment for adequate diffusion to take place. In the particular case of Malo kingi, its small size (less than that of an average human thumbnail) helps to even further increase its surface area to volume ratio.
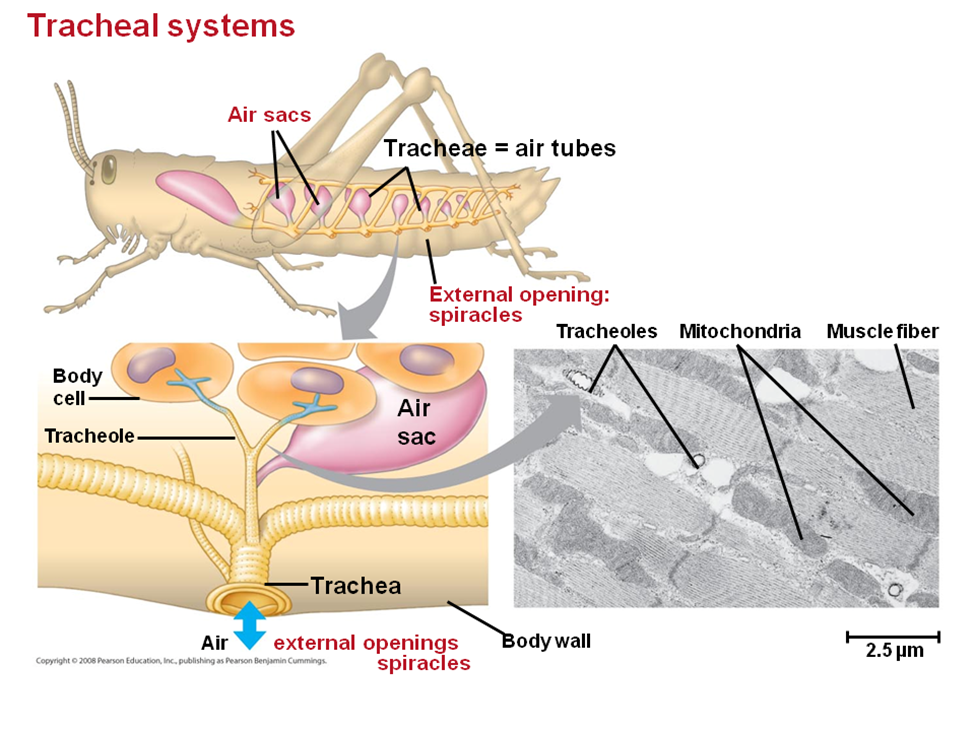
Tracheal System – Grasshopper
Most terrestrial organisms in the uniramia subphylum of arthropods (insects, centipedes, millipedes, and other myriapods) utilize a structurally simple system for gas exchange called a tracheal system. The trachea is made up of air-filled internal tubes with finer branches called tracheoles extending to all parts of the organism's body. A tracheal system carries oxygen directly to its site of use: the body's cells. It opens to the outside environment at small holes on the insect's body, called spiracles. Usually, an insect will have two pairs of spiracles on its thorax and seven or eight pairs on its abdomen. Although structurally simple, tracheal systems are impressively effective in the bodies of insects, allowing for the highest rate of gas exchange relative to body mass out of any animals. Rather than using a diaphragm like a human, a terrestrial insect, such as a grasshopper, expands and contracts the entirety of its abdomen, which changes its tracheal diameter, moving air in and out. During inspiration, air diffuses into fluid-filled tips at the ends of the tracheoles, then oxygen gas, dissolved in the fluid, diffuses into the surrounding cells. Carbon dioxide diffuses from the cells to the tips of the tracheoles, then it can be diffused out directly through skin or it can exit through the spiracles.
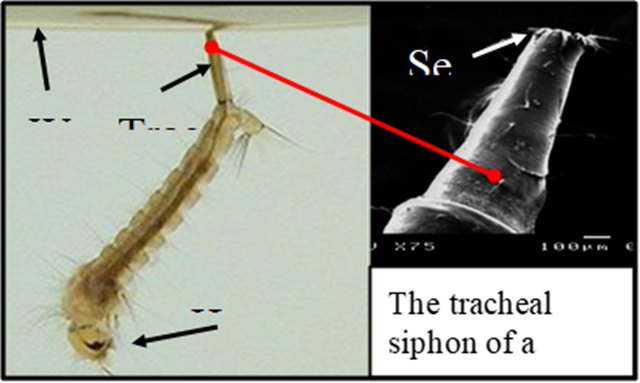
Tracheal Siphon – Mosquito Larva
A tracheal siphon is a hollow tube-like structure used by some aquatic insects that still derive their source of oxygen from the atmosphere, rather than the surrounding water. Often this is necessary for organisms living in oxygen deficient water. Usually this is warm, stagnant, or polluted water where large amounts of bacteria consume considerable quantities of the water's dissolved oxygen. Tracheal siphons work on the same principle as a diver's snorkel. In mosquito larvae, for example, the siphon tube is an extension of the posterior spiracles. An opening at the end of the siphon is guarded by a ring of closely spaced hairs with a hydrophobic, non-polar, waterproof coating. At the air-water interface, these hairs break the surface tension of the water and maintain an open airway. When the insect dives, water pressure pushes the hairs close together so they seal off the opening and keep water out. A siphon mechanism capable of opening and then closing such that it is watertight, using nothing but hairs, is only possible on a small scale, where the effects of surface tension are experienced to a relatively much greater degree than on a larger scale, where the effects can often be negligible.
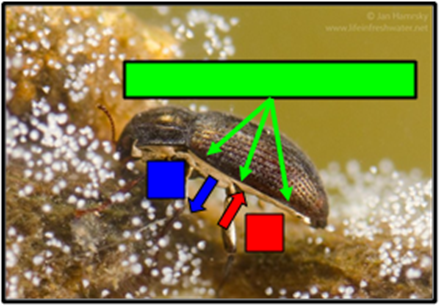
Plastron – Riffle Beetle
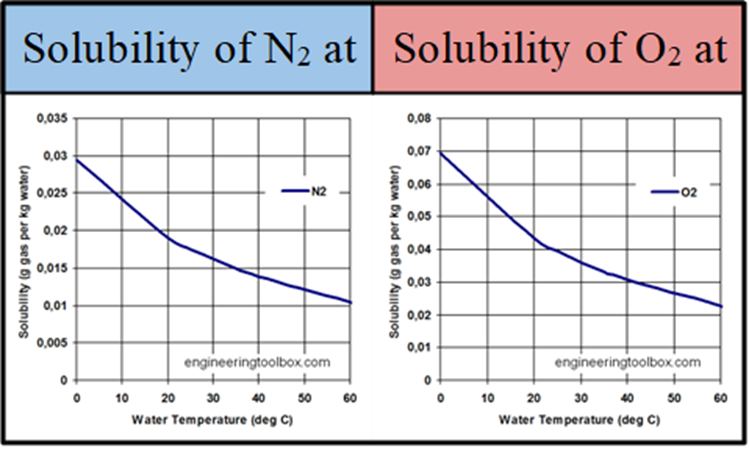
The ventral surface of the body of adult riffle beetles is covered with a layer of hydrophobic hairs called setae. The density of this layer can reach up to several million hairs per square-millimeter. When the beetle dives under water, the hairs trap a thin layer of air against its body called a plastron. Riffle beetles can use this plastron as a physical gill. Since the hairs keep the plastron's volume constant, as the insect consumes oxygen, a pressure difference is created, which results in more oxygen diffusing into the plastron from the water. Additionally, nitrogen gas gradually diffuses out of the plastron, and since the concentration of dissolved nitrogen in the water is quite low, some of the lost nitrogen is replaced by more oxygen, which is more prevalent due to its higher solubility potential in water.
Oxygen gas that is inside the plastron enters the beetle's tracheal system through spiracles, then progresses to the rest of the body just like with a typical terrestrial insect such as a grasshopper. Insects that have plastrons like riffle beetles are capable of spending effectively indefinite amounts of time underwater, making it an advantageous adaptation for an aquatic lifestyle, while also still allowing for normal respiratory function in a terrestrial environment.
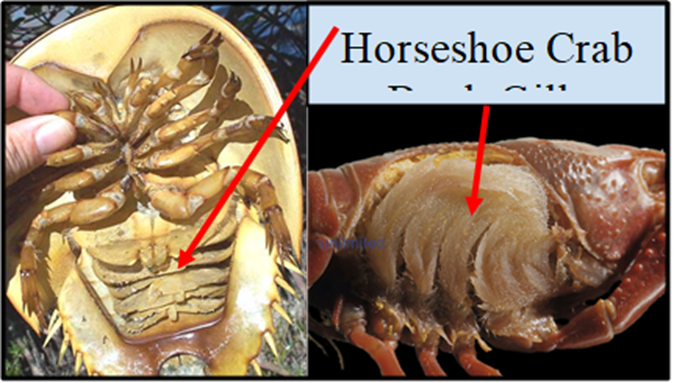
Book Gills – Hermit Crab
Book gills are used in various crustaceans and in horseshoe crabs, which are actually more closely related to arachnids. They are often flap-like or crescentic-shaped in appearance and tend to be located on the undersides of arthropods. A book gill is made up of several thin structures called gill lamellae or gill plates, which are connected to the gill base at a right angle, similar to how the pages of a book are connected to the book spine. The book gills are bathed with water and supplied with copious hemolymph, which is a fluid in arthropods that is analogous to blood in vertebrates. With the assistance of the heart, deoxygenated hemolymph is carried to the gill lamellae from the body's cells, where it is oxygenated and CO2 is removed. The hemolymph is then returned back to the body's cells. The thin membranes of the gill lamellae have a very small diffusion distance between the water and the hemolymph. Additionally, the gill lamellae help increase the surface area of the respiratory surface, optimizing the organism's ability to undergo gas exchange. Muscle movements in the arthropod help to cycle water through the book gills so that the concentration of oxygen is higher and the concentration of carbon dioxide is lower outside the gills than in the gills' hemolymph. This is essentially the same reason that humans breathe.
One particularly interesting example is the hermit crab. Hermit crabs, despite having book gills, can stay in a terrestrial environment for weeks at a time. Most crabs are actually capable of storing some water inside the carapace, on the book gills. If adequate moisture is present on the gill lamellae, then gases can diffuse from the air, to the moisture, to the hemolymph, and vice versa. Therefore, crabs are capable of respiration outside water as long as the book gills remain sufficiently moist. In some ways, this mechanism bears similarities to the plastron used by some aquatic insects. A plastron uses air stored around the respiratory system to allow an “air-breathing” animal to survive underwater, whereas a crab uses water stored around its “water-breathing” respiratory system to survive out in a terrestrial environment. In the particular case of hermit crabs, additional water can be stored inside the shell, which can be used to bathe the gills for an extended period of time.
Book Lungs – Spider
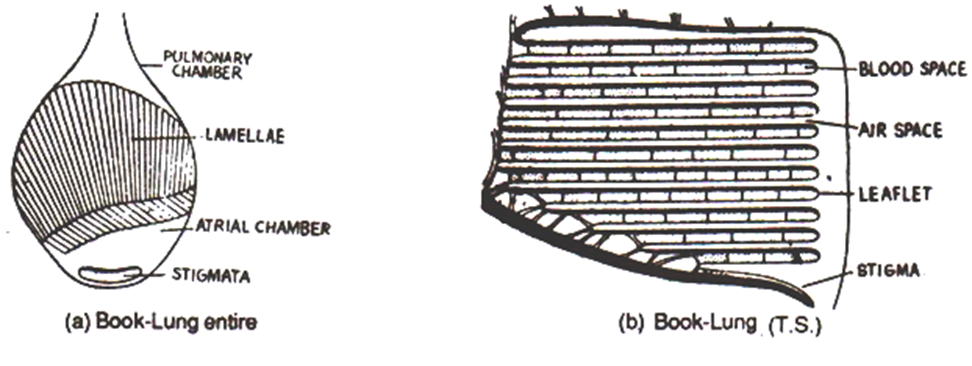
Book lungs are essentially internalized book gills and are found in some terrestrial arachnids, such as spiders and scorpions. Each book lung consists of a pocket partially occupied by a highly folded inner lining where gases diffuse between the air and the hemolymph. A book lung is separated into two sections or chambers: a dorsal chamber and a ventral chamber. The dorsal (pulmonary) chamber contains approximately one hundred fifty leaf-like lamellae (leaflets), formed by the in-folding of the ventral abdominal wall. The lamellae are stacked upon each other in a way reminiscent of the pages in a book, hence the name “book lung.” Each lamellae is continuously supplied a significant quantity of hemolymph, just like how a human lung is constantly supplied large amounts of blood. The ventral (atrial) chamber is not folded and only contains air. Arachnids, such as a typical garden spider, control the movement of air in and out of the book lungs by atrial and dorso-ventral muscles. When the muscles in the abdomen contract, the book lung compresses, forcing air out of the dorsal chamber, into the ventral chamber, and out through a specific type of spiracle called a stigma. When the muscles are relaxed, the book lung regains its shape, which causes air to enter through the stigma, due to the lower pressure inside. Spiders have one or two pairs of book lungs, which are always positioned next to each other on the underside of the abdomen. Scorpions have four pairs of book lungs, which are found along the entire underside of the body.
Book lungs are believed to have evolutionarily developed from book gills when the ancestors of arachnids began to live on land. Additionally, it is possible that the tracheal systems found in opiliones (harvestmen) evolved from book lungs since some spiders do not have leaflets in their lung cavities, but rather, they perform gas exchange with the wall of the chamber itself and use narrow, tube-like structures that branch off the main chamber to increase surface area. The opilione tracheal system is believed to be merely an analogous structure to the insect tracheal system. Insect trachea are believed to have developed independently of arachnid book lungs since insects did not originate from terrestrial arachnids.
The Extraordinary Respiratory System of the Axolotl
Axolotls are among the most unique vertebrates in the world since they possess many unusual physical traits such as the ability to accept partial brain transplants from other axolotls and the ability to regenerate entire limbs. Axolotls are just as unique in terms of their methods of gas exchange too. Unlike humans, who only have one method of gas exchange, an axolotl can undergo gas exchange by four different methods. Firstly, an axolotl can use its three pairs of iconic external gills, called gill rami. External gills are a very rare structure among adult vertebrates and are only common among amphibian larvae and various arthropod larvae. There are a few potential drawbacks associated with external gill structures such as the fact that they cause more drag than internal gills, can easily be damaged, and dry out faster. The second of these three issues is mitigated by the axolotl's regenerative abilities, and the third is solved by the fact that the axolotl spends its entire life in an aquatic environment. The gill rami are lined with a large number of filaments called fimbriae, which considerably increase the surface area. Additionally, the membranes of the gill rami are quite thin, resulting in a very small water-blood diffusion distance. The gill rami are also quite vascular as seen by the defining red hue on individuals with low pigmentation. Gill rami maximize respiratory surface area, minimize the water-blood diffusion distance, and have an excellent blood supply, making them a very effective gas exchange interface.

As with other amphibians, axolotls are capable of using their own skin as a respiratory surface by undergoing cutaneous respiration. Axolotls have thin, delicate skin, which allows gas to diffuse through without much hindrance. Some of the oxygen diffuses directly into the skin cells for use in cellular respiration while the rest of it diffuses through to the blood to be transported through the circulatory system to cells in other parts of the body. Carbon dioxide is also expelled out of the body through the skin. The axolotl's circulatory system typically runs blood vessels that have already travelled to many of the body's cells just under the skin. This means that the blood brought here already contains a high concentration of CO2 and a low concentration of low O2, which establishes a concentration gradient and allows diffusion to occur. Additionally, axolotls coat their skin in a layer of mucus which aids in diffusion through the skin and retains moisture.
Axolotls can simultaneously perform a third form of gas exchange, which is buccal respiration. This is carried out through the buccopharyngeal membrane in a similar way to cutaneous respiration. The buccopharyngeal membrane is a highly vascular, thin-walled tissue at the hack of the throat and along the top of the inside of the mouth. The surrounding area in the back of the throat is called the buccal pouch. Axolotls can pulsate the buccal pouch and buccopharyngeal membrane causing water to flow through the oral cavity and diffuse oxygen into the blood. Frogs and toads also use buccal respiration and have much more visible buccal pouches. In these organisms, buccal respiration differs slightly from in axolotls. Frogs and toads inhale air instead of water when they pulsate their buccal pouches.

Finally, axolotls also possess the ability to breathe air through lungs, despite never becoming terrestrial as adults. Breathing through the lungs is the least efficient of the axolotl's various respiratory mechanisms in terms of blood oxygenation rate. Axolotls' lungs are small, simple, and sac-like. The lungs contain folded sections of thin membranes, encircled by capillaries, which resemble the alveoli found in the lungs of reptiles and mammals. The tissues inside the lungs are moist and have a small air-blood diffusion distance, which allows oxygen to enter the capillaries, where it is brought back to the heart by veins, in order to then be pumped around the body. Some CO2 diffuses from the blood into the lungs where it is exhaled, but most of it is still released cutaneously. The axolotl typically only employs its lungs when the oxygen content in the water is too low. Increasing urbanization in and around Mexico City, has caused the water in many parts of the axolotl's natural habitat to become oxygen deficient, due to an increased presence of bacteria and algal blooms resulting from pollution. The ancestors of the axolotl were conventional amphibians which dwelled on land as adults and used their lungs to breathe, which is the primary reason why the axolotl has lungs, despite being highly aquatic.
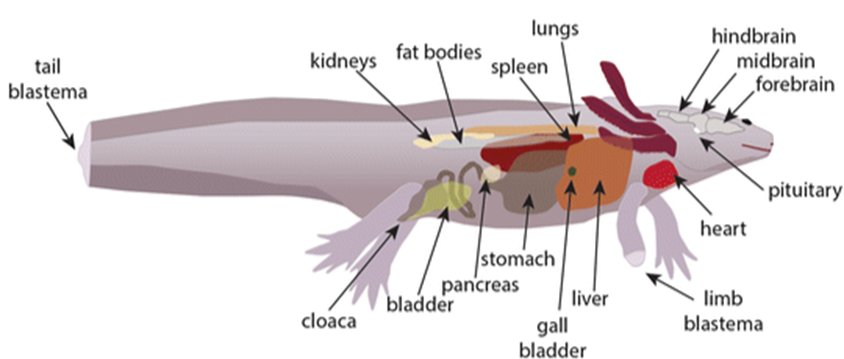
The axolotl is quite a unique creature, which begs the question of how such an unusual organism could have evolved. As mentioned earlier, the axolotl exhibits characteristics in adulthood that typically only appear in the early stages of other amphibians' life cycles, such as external gills and a caudal fin (tail fin), since it does not undergo metamorphosis. This phenomenon is referred to as neoteny. Neoteny is the retention of some immature or larval characteristics into the sexual maturity of an organism. For example, humans exhibit some neotenic primate traits, such as the adulthood retention of short arms and flat faces. The axolotl's habitat may help to explain how it developed to become neotenic. Since the larval stages of amphibians are well adapted to aquatic environments, it is advantageous for the axolotl to retain these characteristics into adulthood because unlike many amphibians, it remains in an aquatic environment its entire life. Natural selection would suggest that in an aquatic environment, the ancestors of the axolotl which exhibited the most larval characteristics later in life had the greatest chance of surviving to sexual maturity and successfully reproducing, eventually resulting in the modern axolotl and its complete absence of metamorphosis.
Lungs – Mammals
Relative to most other organisms, mammals exhibit many traits that require them to have an efficient gas exchange mechanism: they are relatively large, have complex nervous systems, are warm blooded, are often covered with hair or fur, and usually live very active lifestyles. It is for all these reasons that mammals must use highly developed lungs. The lungs used by mammals and reptiles are full of small, highly folded sections of membrane called alveoli. The alveoli typically have walls that are only one-cell-thick, and are surrounded by capillaries on nearly all sides. As a result, lungs have a massive surface area to volume ratio, have a small air-blood diffusion distance, and are highly vascular, all of which makes them an efficient gas exchange mechanism. Mammals live in a wide variety of climates and environments, so cutaneous respiration is not a viable mechanism, since it would be highly impractical to maintain moist skin, especially considering the fact that most mammals are almost completely covered in hair or fur. Lungs are entirely internalized which is another reason why they are an excellent respiratory mechanism for mammals.

Avian Respiratory System
Due to the high amount of energy that must be expended to fly, birds must be capable of extremely high metabolic rates. In turn, this means that birds require large amounts of oxygen and produce large amounts of carbon dioxide relative to their body mass. To accomplish this, birds have developed the most effective lung-based respiratory system on Earth. The lungs of a bird are fairly rigid structures that do not expand and contract the way that the lungs of mammals, reptiles and many amphibians do. Rather, a bird expands and contracts separate structures distributed throughout its body called air sacs in order to ventilate its lungs. The air sacs move air unidirectionally through narrow tubes called parabronchi, which are located inside the rigid lungs.
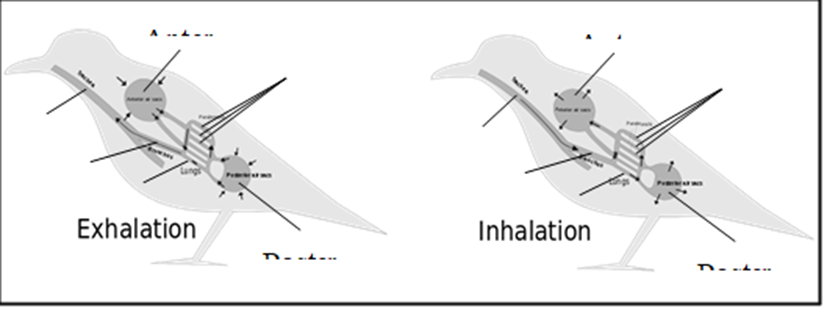
Bird lungs are smaller than those of mammals of comparable size, meaning that their respiratory surface is smaller. Birds make up for this by having the ability to expose their respiratory surface to considerably more air with any given breath. This is because the air sacs account for approximately fifteen percent of a bird's total body volume, whereas the alveoli, which act as the bellows in a mammal's lungs, constitute only about seven percent of the total body volume.

Bibliography
Abraczinskas, C. (2008). File:Cranial sinus and postcranial air sac systems in birds.svg. Wikimedia Commons. Retrieved October 22, 2020, from https://commons.wikimedia.org/wiki/File:Cranial_sinus_and_postcranial_air_sac_systems_in_birds.svg
AquaTerra-Life. (2015, May 27). Macrography: External Gills (Axolotl) || Makrofotographie: Außenkiemen (Ambystoma mexicanum) Youtube. https://www.youtube.com/watch?v=aeejOPlXr-o
Axolotl Nerd. (2019, November 18). How do Axolotls Breathe? Retrieved October 22, 2020, from https://axolotlnerd.com/how-do-axolotls-breathe/
Brown, L. (2016, May 2). Axolotls Four Method of Breathing. https://lollybrown.com/axolotls-four-method-breathing/
Carter, S. (n.d.). Redox Reactions. Lumen Learning. Retrieved October 22, 2020, from https://courses.lumenlearning.com/suny-wmopen-biology1/chapter/redox-reactions/
Charalampidis, C., Youroukou, A., Lazaridis, G., Baka, S., Mpoukovinas, I., Karavasilis, V., Kioumis, I., Pitsiou, G., Papaiwannou, A., Karavergou, A., Tsakiridis, K., Katsikogiannis, N., Sarika, E., Kapanidis, K., Sakkas, L., Korantzis, I., Lampaki, S., Zarogoulidis, K., & Zarogoulidis, P. (2015). Pleura space anatomy. J Thorac Dis, 7(Suppl 1), S27-32. https://doi.org/10.3978/j.issn.2072-1439.2015.01.48
Conrad, J. (2005, December 15). Horseshoe Crabs. Backyard Nature. https://www.backyardnature.net/yucatan/hs_crab.htm
Crew, B. (2013, October 7). The Smallest and Deadliest Kingslayer in the World. Scientific American. https://blogs.scientificamerican.com/running-ponies/the-smallest-and-deadliest-kingslayer-in-the-world/
Cunningham, J. D. (2009, November 17). Crayfish gills. Visuals Unlimited. Retrieved October 22, 2020, from https://visualsunlimited.photoshelter.com/galleries/I0000lFnsL0xQetw/Crayfish-gills
Dubuc, B. (n.d.). The Origin of Mitochondria. The Brain From Top to Bottom. Retrieved October 22, 2020, from https://thebrain.mcgill.ca/flash/a/a_05/a_05_cl/a_05_cl_her/a_05_cl_her.html
Fautin, D. G. (2020, October 20). cnidarian. Encyclopedia Britannica. Retrieved October 22, 2020, from https://www.britannica.com/animal/cnidarian
Feliz, D. (2013, October 10). Ensatina. Elkhorn Slough. https://www.elkhornslough.org/ensatina/
Fish gill. (2020, October 5). Wikipedia. Retrieved October 22, 2020, from https://en.wikipedia.org/w/index.php?title=Fish_gill&oldid=981979269
Gaillard, F. (n.d.). Normal chest x-ray. Radiopaedia. Retrieved October 22, 2020, from https://radiopaedia.org/cases/normal-chest-x-ray
Gould, S. E. (2012, July 29). The origin of breathing: how bacteria learnt to use oxygen. Scientific American. https://blogs.scientificamerican.com/lab-rat/the-origin-of-breathing-how-bacteria-learnt-to-use-oxygen/
Graham, J. B. (2011). Air-Breathing Fishes | Respiratory Adaptations for Air-Breathing Fishes. In A. P. Farrell (Ed.), Encyclopedia of Fish Physiology (pp. 1861-1874). Academic Press. https://doi.org/https://doi.org/10.1016/B978-0-12-374553-8.00045-9
Hammond, G. (2009). Elmidae. Animal Diversity Web. Retrieved October 22, 2020, from https://animaldiversity.org/accounts/Elmidae/
Hsia, C. C., Schmitz, A., Lambertz, M., Perry, S. F., & Maina, J. N. (2013). Evolution of air breathing: oxygen homeostasis and the transitions from water to land and sky. Compr Physiol, 3(2), 849-915. https://doi.org/10.1002/cphy.c120003
Hughes, G. M. (1966). The dimensions of fish gills in relation to their function. Journal of Experimental Biology, 45(1), 177-195.
Hughes, G. M., & Grimstone, A. V. (1965). The fine structure of the secondary lamellae of the gills of Gadus pollachius. Journal of Cell Science, s3-106(76), 343-353. https://doi.org/10.1242/jcs.s3-106.76.343
Hughes, G. M., Perry, S. F., & Piiper, J. (1986). Morphometry of the gills of the elasmobranch Scyliorhinus stellaris in relation to body size. Journal of Experimental Biology, 121(1), 27-42.
Kingsley, C. (n.d.). Lung Capacity: What You Need To Know. Lung Institute. Retrieved October 22, 2020, from https://lunginstitute.com/lung-capacity/
Levitzky, M. G. (2017). Pulmonary Physiology (9th ed.). McGraw-Hill Education.
Li, G., Lv, X., Zhou, J., Shen, C., Xia, D., Xie, H., & Luo, Y. (2018). Are the surface areas of the gills and body involved with changing metabolic scaling with temperature? Journal of Experimental Biology, 221(8). https://doi.org/10.1242/jeb.174474
Lungs. (2017, September 2). The Respiratory System. https://www.therespiratorysystem.com/lungs/
Madathilparambil, S., Yu, B., Machado-Aranda, D., Bender, M., Ochoa-Frongia, L., Helinski, J., Davidson, B., Knight, P., Hogaboam, C., Moore, B., & Raghavendran, K. (2012). Role of Macrophage Chemoattractant Protein-1 in Acute Inflammation after Lung Contusion. American journal of respiratory cell and molecular biology, 46, 797-806. https://doi.org/10.1165/rcmb.2011-0358OC
Maina, J. N. (2002). Structure, function and evolution of the gas exchangers: comparative perspectives. J Anat, 201(4), 281-304. https://doi.org/10.1046/j.1469-7580.2002.00099.x
Metzger, R. J., Klein, O. D., Martin, G. R., & Krasnow, M. A. (2008). The branching programme of mouse lung development. Nature, 453(7196), 745-750. https://doi.org/10.1038/nature07005
Mexican Axolotls: Characteristics, reproduction, hábitats and more. (n.d.). Our Marine Species. Retrieved October 22, 2020, from http://ourmarinespecies.com/c-other-species/mexican-axolotls/
Meyer, J. (2016, March 25). Respiration in Aquatic Insects. General Entomology. Retrieved October 22, 2020, from https://projects.ncsu.edu/cals/course/ent425/library/tutorials/internal_anatomy/aquatic_insects.html
Molnar, C., & Gair, J. (2015). 20.1 Systems of Gas Exchange. In Concepts of Biology – 1st Canadian Edition. BCcampus. https://opentextbc.ca/biology/chapter/20-1-systems-of-gas-exchange/
Molnar, C., & Gair, J. (2015). 20.2 Gas Exchange across Respiratory Surfaces. In Concepts of Biology – 1st Canadian Edition. BCcampus. https://opentextbc.ca/biology/chapter/20-2-gas-exchange-across-respiratory-surfaces/
Needham, D. (2012). Axolotl Canada,. Axolotl Canada. Retrieved October 22, 2020, from http://axolotlcanada.com/page11/page11.html
O'Rourke, D. P., & Jenkins, A. L. (2008). Chapter 20 – Anesthesia and Analgesia in Amphibians. In R. E. Fish, M. J. Brown, P. J. Danneman, & A. Z. Karas (Eds.), Anesthesia and Analgesia in Laboratory Animals (Second Edition) (pp. 511-518). Academic Press. https://doi.org/https://doi.org/10.1016/B978-012373898-1.50024-3
Ostadfar, A. (2016). Chapter 1 – Fluid Mechanics and Biofluids Principles. In A. Ostadfar (Ed.), Biofluid Mechanics (pp. 1-60). Academic Press. https://doi.org/https://doi.org/10.1016/B978-0-12-802408-9.00001-6
Palzenberger, M., & Pohla, H. (1992). Gill surface area of water-breathing freshwater fish. Reviews in Fish Biology and Fisheries, 2(3), 187-216. https://doi.org/10.1007/BF00045037
Park, K., Kim, W., & Kim, H. Y. (2014). Optimal lamellar arrangement in fish gills. Proc Natl Acad Sci U S A, 111(22), 8067-8070. https://doi.org/10.1073/pnas.1403621111
Powers, K. A., & Dhamoon, A. S. (2020). Physiology, Pulmonary Ventilation and Perfusion. In StatPearls [Internet]. StatPearls Publishing. https://www.ncbi.nlm.nih.gov/books/NBK539907/
Rodenbaugh, D. W., Collins, H. L., & Dicarlo, S. E. (2003). A simple model for understanding cohesive forces of the intrapleural space. Advances in Physiology Education, 27(1), 42-43. https://doi.org/10.1152/advan.00058.2002
Scitable. (2014). Cell Energy and Cell Functions. Nature Education. Retrieved October 22, 2020, from https://www.nature.com/scitable/topicpage/cell-energy-and-cell-functions-14024533/
Shyamal, L. (2007, December). File:BirdRespiration.svg. Wikimedia Commons. Retrieved October 22, 2020, from https://commons.wikimedia.org/wiki/File:BirdRespiration.svg
Singh, B., Yadav, A., Ojha, J., & Munshi, J. D. (1981). Gross structure and dimensions of the gills of an intestinal airbreathing fish (Lepidocephalichthys guntea). Copeia, 1981(1), 224-229.
Soil Biology. (2018, March 9). CharGrow. https://char-grow.com/soil-biology
Speller, J. (2020). Gas Exchange. TeachMe Physiology. Retrieved October 22, 2020, from https://teachmephysiology.com/respiratory-system/gas-exchange/gas-exchange/
Sultan, F. (2018). Relationship between total length and gill surface area in orange spotted grouper, epinephelus coioides (Hamilton, 1822) The Iraqi Journal of Agricultural Sciences, 49, 268-268.
Systems of Gas Exchange. (n.d.). Course Hero. Retrieved October 22, 2020, from https://www.coursehero.com/study-guides/boundless-biology/systems-of-gas-exchange/
Taylor. (n.d.). How Long Can Hermit Crabs Go Without Food, Water, & Proper Care? Hermit Crab Owner. Retrieved October 22, 2020, from https://hermitcrabowner.com/how-long-can-hermit-crabs-go-without-food-water-proper-care/
ToolBox, E. (2008). Solubility of Gases in Water vs. Temperature. Retrieved October 22, 2020, from https://www.engineeringtoolbox.com/gases-solubility-water-d_1148.html
Vandana S. (n.d.). Book Lung of Scorpion (With Diagram) | Zoology. Biology Discussion. Retrieved October 22, 2020, from https://www.biologydiscussion.com/zoology/practicals/book-lung-of-scorpion-with-diagram-zoology/60387
Zoological Society of Assam. (2020, May 18). Respiration in Arthropoda, YouTube. https://www.youtube.com/watch?v=2Zp0_pJ-edE Zug, G. R. (2006). Amphibians. Encyclopedia Britannica. Retrieved October 22, 2020, from https://www.britannica.com/science/respiratory-system/Amphibians