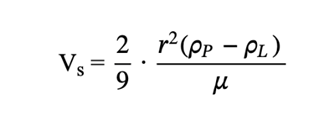




ABSTRACT
The intricate silica skeletons of Radiolaria, a type of marine microorganism, exhibit striking optical fiber-like properties, offering a potential roadmap for future innovations in the optical field. Beyond their applications for photonics, radiolarians are fascinating models for studying buoyancy control. They exhibit a variety of adaptive mechanisms that alter their specific gravity, such as spine shedding, skeleton and shell secretion, complex ion pumping, and pseudopodia extension. These eukaryotes also stand out from an architectural point of view, due to the captivating glass-like spherical skeletons characterised, for some, with geometrical tessellation. Moreover, radiolarite, the fossilized remains of radiolarians, provides an extensive archive for geohistorical research. The distributions of these microfossils serve as a record for past geological events, such as tectonic shifts, volcanic activity, and hydrothermal events. Collectively, these insights not only highlight fundamental principles in biology, geology, and physics, but also point towards a horizon of interdisciplinary innovations.
INTRODUCTION
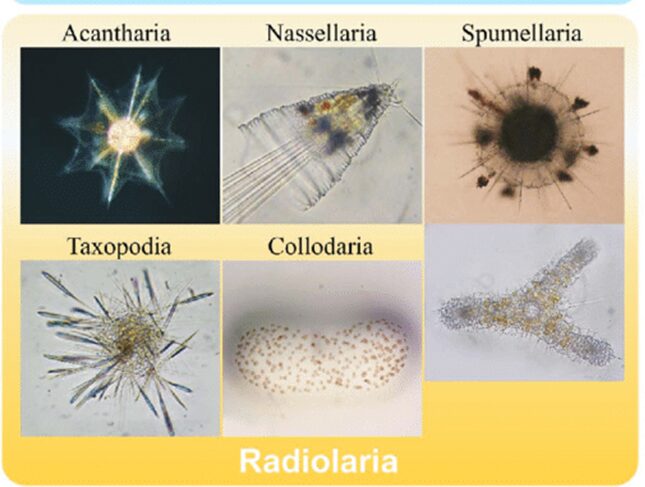
Amongst the diversity of oceanic species and realms in the oceans, protists such as Radiolaria are a diverse group of unicellular eukaryotes that have raised interest not only within the scientific community but also in architecture and arts due to their intricate mineral skeletons (Jungck et al., 2019). Averaging from tenths to hundreds of millimeters in size, Radiolaria are classified into 6 different orders according to their chemical composition and structure of their skeleton (Matul & Mohan, 2017; Biard, 2022). The six orders of classification of Radiolaria are the Acantharia and the Taxopodida as well as the polycystine radiolarians, which are composed of the Spumellaria, the Nassellaria, the Collodaria and the Orodaria (Fig.1).
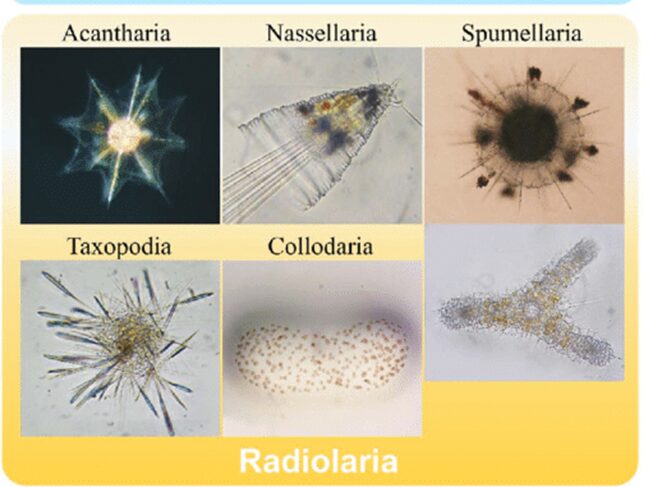
Fig. 1 Visual of the different classifications of Radiolaria (Nakamura et al., 2019)
The top and bottom left are Acantharia and Taxopodia respectively. The top and bottom in the middle are Nassellaria and Collodaria, respectively. Lastly, the top and bottom right are Spumellaria and Orodaria.
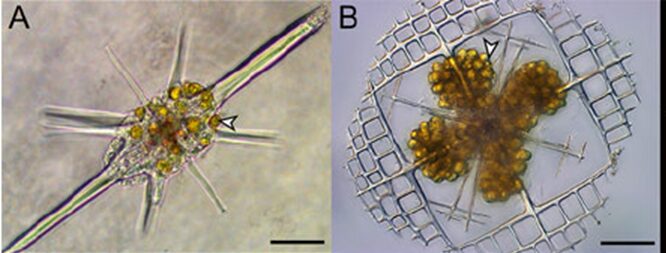
These different orders biomineralize different minerals like strontium sulfate (SrSO4) and opaline silica (SiO2nH2O) (Biard, 2022). Radiolarians also captivated scientists due to their skeleton remains that serve an important purpose after death, as they sink to the bottom of the ocean and incorporate into the sediments (Biard, 2022). This allows for scientists to have an extensive fossil record for radiolarians. Yet, these types of records do not necessarily help the scientific community extend their knowledge and understanding on radiolarian ecology. In fact, sampling living Radiolaria to study them proved to be challenging for scientists. However, biotic interactions between them and photosymbionts have been observed. This interaction is also known as photosymbiosis, where a host organism, such as Radiolaria, have microalgae, the photosymbionts, living in their cells (Fig. 2). In turn, this interaction benefits both species; the host is provided with products from the photosynthetic activities of the microalgae, while the latter lives in an environment that provides nutriments and resources (Decelle et al., 2012). The specific microalgae that Radiolaria are known to host are dinoflagellates, prasinophytes or haptophytes. In particular, the Acantharean order of Radiolaria are known to host such symbiotic microalgae, thus explaining why its population roams more in surface waters compared to the other orders of Radiolaria (Decelle et Al., 2012). Radiolaria specifically benefits from carbon-rich products from the photosynthesis carried out by these photosymbionts, but in this case, it is suggested that it may not be equally favorable for both species, advantaging the host to a greater extent (Biard, 2022).
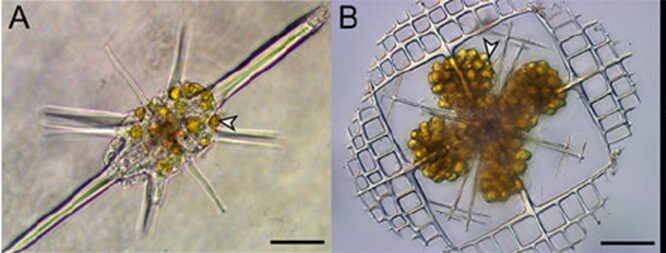
Fig. 2 Photosymbiotic interaction between Acantharia and the photosymbionts Phaeocystis (Haptophytes), indicated with arrows (Decelle et al., 2015).
There do not seem to be distinctive predators and preys related to Radiolaria, which prevents scientists from considering them in marine food webs, because their role in it is not clear. However, it is known that most of them are phagotrophs, feeding on various types of prey, like bacteria, dinoflagellates, protists, and multicellular heterotrophs by ingesting them using food vacuoles (Biard, 2022; Fenchel, 2013). It is also hypothesized that the rest of the radiolarians that are not phagotrophs are in fact detritivores, consuming inorganic matter and decomposing organisms (Biard, 2022; Schowalter, 2016). Furthermore, scientists have identified potential predators of Radiolaria as crustacean species such as amphipods, decapods, euphausids, isopods, ostracods, mysids, and copepods, as well as molluscs, annelids, gelatinous plankton and fishes like adults and larvae (Biard, 2022).
Concerning the vertical distribution of radiolarian populations in the tropical and subtropical waters, polycystine radiolarians are mostly concentrated in the upper layers, such as 0 to 100 meters deep. The population density then gradually decreases at deeper ocean depths (Boltovskoy, 2017). As for the cold waters of the northern hemisphere, the population density is overall lower, but more concentrated in shallower waters, which is similar for the southern polar and sub polar waters, apart from the fact that they are more densely populated than the northern ones (Boltovskoy, 2017). All in all, radiolarian populations seem to thrive in waters with higher temperatures and at an average sea depth of 200 meters (Matul & Mohan, 2017). Furthermore, the study of Radiolarian ecology and their roles in biochemical cycles is key to understanding ocean geology, history, marine evolution, and climate change (Sharma & Daneshian, 1998).
SKELETAL STRUCTURE
Structural adaptations for the different classifications
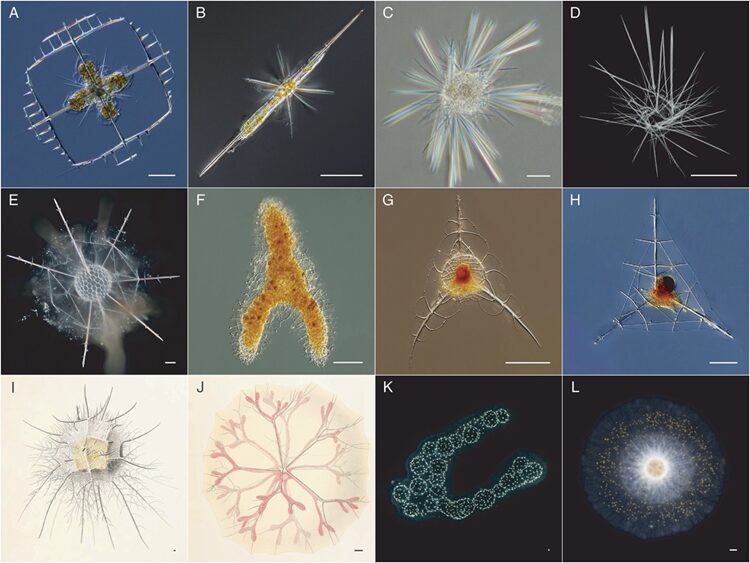
In general, radiolarian skeletons are composed of siliceous shells with a thickness ranging from 10 to 100 μm and an internal cavity with a diameter around 30 to 250 μm. The internal part may be filled with calcite, pyrite, siliceous particles or organic matter that form multiple pores in the cavity, thus resembling honeycomb, that, when fully developed, allows the Radiolaria to withstand a lot of pressure due the structure’s compressive strength (Zhou et al., 2022). Nevertheless, not all radiolarian skeletons are structurally the same, so classification of Radiolaria is determined according to the chemical composition of their skeletons as well as its structural components. For the Acantharean order, they biomineralize strontium sulfate (SrO4), while all the other classes, which are part of the Taxopodida and the Polycystines, have skeletons mineralized with opaline silica (SiO2 nH2O) (Biard, 2022). Acantharia also differ in the development of their skeletons because their structure starts developing from the inside of the central capsule, compared to all the other orders where it develops from the outside (Afanasieva & Amon, 2014). Since they biomineralize strontium sulfate (SrO4), which is highly undersaturated in the ocean, their skeleton dissolves when they die, contrary to the skeletons of Polycystines and Taxopodida that become buried in the seabed and become fossils (Zhou et al., 2022). The structure of the Acantharia skeleton is also unique because of its regularity and predictability (Fig.3, A-B). Their spines are arranged in an organized pattern where they intersect to a central point at 90° angles, which demonstrates a simpler geometry compared to the more complicated skeleton structures of other Radiolaria (Anderson, 1981).
Other types of radiolarian skeletons are more intricate and spheroidal in shape with better space for cytoplasmic matter. The central point is not an intersection for their spines, rather the location of their nuclei. In contrast, Acantharean structures are characterized by long spines emerging from their spherical shells. The latter are further reenforced by a geometric lattice which increases the probability to capture prey due to a better surface area and due to the spines, that emerges from their shells and allows them to anchor themselves while trying to secure an active prey (Anderson, 1981). More specifically, Taxopodida are characterized by their multiple spikes coming out from their central capsule, which are called axopodia and are silica-based (Fig.3, C-D) (Biard, 2022). As for Spumellaria and Nassellaria, they stand out because their silicified skeletons are shaped as spheres or as cones (Fig.3, E-F) (Biard, 2022). Nassellarian species tend to be cone-like in shape, also known as cephalis, that are characterized by a symmetrical axis that allows rotation (Fig.3, G-H) (Anderson, 1981). Orodaria and Collodaria are distinct due to their size, often reaching a size of a thousand micrometers, while the average size for other radiolarians ranges in the hundreds of micrometers (Fig.3, I-L). Collodaria (Fig.3, K-L) are also known to have a skeleton structure that lacks silica and resembles spherical sponges with their very small, silicified spicules (Biard, 2022).
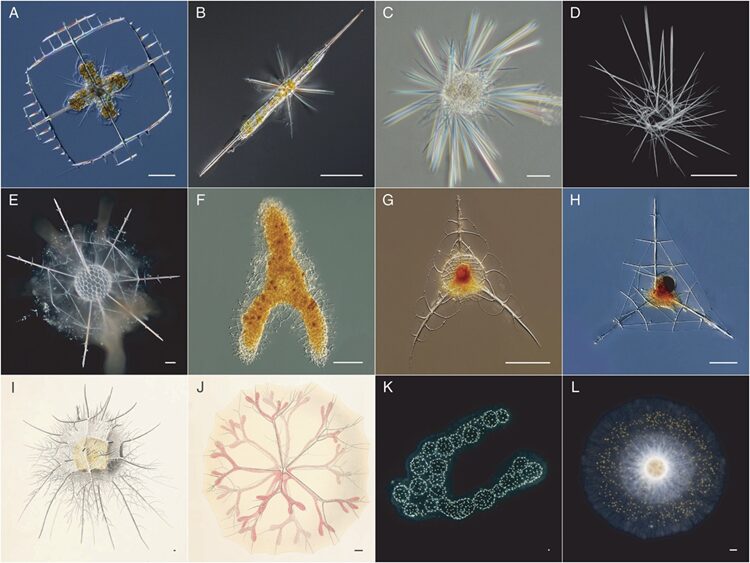
Fig. 3 Observed structures of the different classification of Radiolaria (Biard, 2022).
A-B: Acantharia
C-D: Taxopodida
E-F: Spumellaria (Polycystine)
G-H: Nassellaria (Polycystine)
I-J: Orodaria (Polycystine)
K-L: Collodaria (Polycystine)
Morphogenesis of Silica in Radiolaria
While the formation of the skeleton of Radiolaria can be caused by chemical principles, there is a chance that a large part could also be formed due to physical laws. In fact, it was suggested that the tightly packed vesicles of the extracapsular cytoplasm create a surface tension at the intersections. These intersections formed a hexagonal shaped pattern around each vesicle. These regions of surface tension could attract silica deposition, thus creating multiple hexagonal rings that form a spherical skeleton characterised with a tessellation (Fig.4) (Anderson, 1981). Additionally, according to a quantitative image analysis on radiolarian fossils, most polygon rings from their skeleton tessellation are hexagons, although there are still some cases of pentagons rings and rare cases of quadrilaterals, all of which are mostly composed of trigonal junctions (Fig. 5) (Jungck et al., 2019). However, upon more electron microscopic research, the silica deposition seems more likely to be guided by cytokalymma synthesis on the skeleton. More precisely, the skeleton of Radiolaria is shown to be covered by a layer of cytoplasm, corresponding to the cytokalymma. When evaluating the skeleton in sections, the thinnest skeletal parts had a thinner layer of cytokalamma on the innermost parts and with gradually increasing thickness at the outermost sections. Consequently, this suggests that the deposition of cytokalamma happened before the deposition of silica, thus serving as a mold of matrix for the silica to fill in and form the architecture of the skeleton (Anderson, 1981).

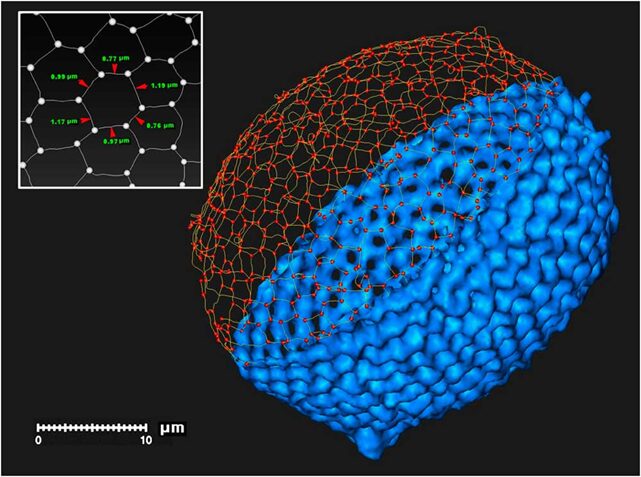
Fig. 4 Photographs of spherical Radiolaria skeleton (Yoshino et al., 2012).
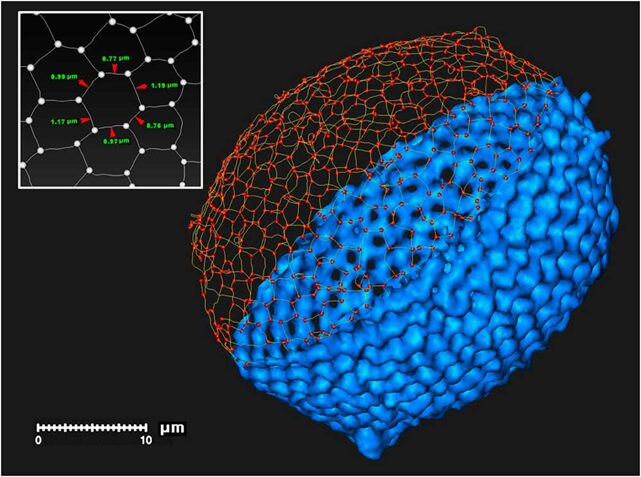
Fig. 5 3D digital modelof a radiolarian skeleton (Wagner et al., 2015).
This digital model illustrates the polygon tessellation of a radiolarian skeleton with the segments and nodes that compose it.
Optical properties of silica
Radiolarians have similar characteristics to sea glass sponges and diatomaceous, such as their silica skeleton having optical properties, like sea glass sponges (Fig. 6). A study has found that these spicules are biological fibers that contain a core with a high refractive index and a surrounding layer with a lower refractive index. Embedded spicules have the properties of single (or few mode) waveguides allowing light to be guided and confined in the core (Sundar et al., 2003). The use of these silica fibers could revolutionize the technology of optical fibers since these spicules can be formed at ambient temperatures whereas commercial optical fibers require high-temperature manufacturing. For example, it could potentially be implemented in the advancement of active optoelectronic devices such as optical fiber amplifiers and lasers. The complexity of the mechanical structure helps with acquiring the strength of the skeleton of these organisms and the shape of their spicules (Kulchin et al., 2008). These spicules can also be treated as photonic crystals. They are composed of many thin layers of organosilicon that produce a pattern in the way they transmit light.

Fig. 6 (left) Spicules(arrow) shown in the silica structure of the glass sponge (Sundar et al., 2003). (right) Photomicrograph of a spicule’s cross-section of a sea glass sponge (Pheronema sp.) (magnification 1.157x) (Kulchin et al., 2008).
MOVING THROUGH WATER
Pseudopodia
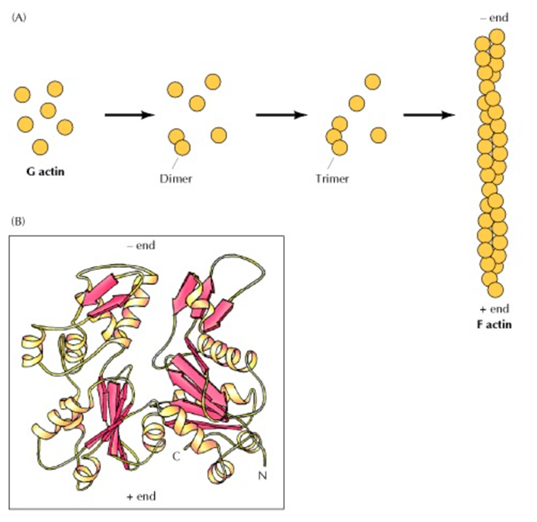
Many microorganisms found on Earth owe their ability to move about their environment to pseudopodia. Pseudopodia are limb-like projections that extend from a microorganism in the direction of movement (Chodniewicz & Klemke, 2004). They are mainly composed of cytoplasmic extensions that are reinforced by tubulin or actomyosin cytoskeletons. (Lara et al., 2020) The use of a cytoplasm base makes it possible to maneuver skillfully, while the cytoskeleton elements provide structural support which enhance the stability of these microorganisms. The projection of these “limbs” is made possible due to the assembly and disassembly of the actin-filaments. (Vicker, 2000) Since actin filament polymerization is reversible, they can polymerize and depolymerize into monomers depending on the ionic character of the host solution. (Cooper, 2000) This process is mediated by many different proteins operating in a signaling pathway that functions as an on/off switch for the polymerization of actin filaments (Vicker, 2002). This assembly, both of actin-filaments and of tubulin, are the driving force behind the locomotion of pseudopodia (Fig.7).
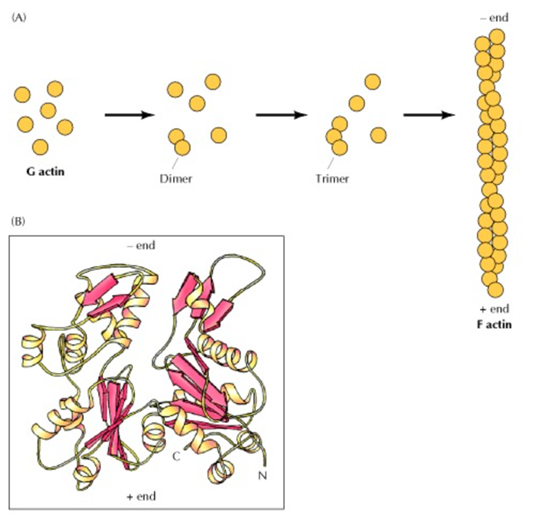
Fig. 7 (A) Polymerization of actin monomers (G actin) to form actin filaments, actin filaments are essentially long chains of actin monomers. (B) A drawing representation of the structure of an actin monomer (Cooper, 2000).
Pseudopodia in radiolarians come in several different types, including axopodia and reticulopodia (Fig.8) (Suzuki & Not, 2015). Axopodia are straight, long pseudopodia whose stabilizing element is an array of microtubules that fill the axoneme (or the axial filament). This type of pseudopodia is a much more complicated structure than others since they have a central core (the axoneme) that’s surrounded by cytoplasm that flows. The axoneme is comprised of a bundle of around 600 microtubules that fit together in a cross-linked pattern (Ishitani, Febvre-Chevalier, & Febvre, 2016; Britannica, 2023). These cross-linked patterns have been observed as branched, hexagonal, and dodecagonal. Since the capture of prey for radiolarians depends on the probability of them running into another organism, having these long pseudopodia allows them to increase their rate of endocytosis of food. This ability is further enhanced by specialized prey-capturing organelles that can be observed on the outer layer of these cytoplasmic extensions (Marshall, 2020).

Fig. 8 Major Radiolaria classifications based on type of pseudopodia extension. As observed, Radiolarian pseudopods can come in many shapes and lengths. The number of pseudopods on a given organisms vary heavily as well (Suzuki & Not, 2015).
The other type of pseudopodia observed on Radiolaria are reticulopodia, which are the main capturers of prey. Reticulopodia are complex network systems without axonemes. Instead, they are made up of thin, fine threads that work as a net. Once another organism gets tangled in the cytoplasmic net, it gets taken by a vacuole into the ectoplasm to be digested. These large nets can increase the probability of a Radiolaria running into a prey organism, which in turn increases their rate of food ingestion. Most Radiolaria are heterotrophic and omnivores, so they get their energy by intaking other organisms. These other organisms include protozoans like diatoms, and even certain crustaceans like copepods (Ishitani et al., 2016).
Just as reticulopodia’s main job is food ingestion, axopodia’s main function is locomotion. In general, protists can move through their environment in three different ways. These include beating of cilia/flagella, rowing of axopods, and crawling (Febvre-Chevalier & Febvre, 1994). In the case of Radiolaria, they primarily rely on the rowing motion of their axopodia. Their axoneme cores make their structure more rigid than cilia or flagella, so Radiolaria are able to use them as oars in the water. Movement of axopodia comes from their base, which are calcium-controlled, contractile microfilaments (Suzuki & Not, 2015). Changes in calcium concentration in the external medium is imperative for the motility of Radiolaria. In fact, it has been observed that a calcium concentration of approximately 0.1 mM is necessary for contraction of pseudopodia’s core (Soden, 2011).
The bundles of stable microtubules that make up the core are connected to the core of the Radiolaria. They employ the assembly and disassembly of actin-filaments to extend and retract from the cell’s skeletal structure. The external filaments facilitate the coordinated movements that make up the motile cycle of the axopodia, which occurs in two steps. The first is the active stroke, in which the axopodia moves forward in the direction of motion. The second is the recovery stroke, where the axopodia retracts and briefly stops before the next motile cycle begins. These mechanics of propagation can be viewed similarly to the rowing of a boat. To move forward, momentum is transferred to water through the pulling back (recovery stroke) of axopodia (Pulman, 2005). The momentum of water causes motion of the Radiolaria in the opposite direction. This is caused by conservation of momentum. So, the momentum transferred to water must equal the momentum of the Radiolaria after the recovery stroke. The results and force cause a forward acceleration. The long shape of axopodia can move a lot of water slowly instead of a little bit of water quickly, which is energetically favourable. There are, of course, draw backs to this type of propagation. The acceleration is opposed by various sources of drag force. These include friction between the surface of the Radiolaria and the water (i.e., fluid drag forces) and wave drag caused by the creation of waves by the transfer of kinetic energy to surrounding water.
Buoyancy Regulation
Radiolaria belong to a group of zooplankton that have relatively heavy skeletons, which makes them much less motile than other types of zooplankton. This means that vertical motion and buoyancy are crucial for Radiolaria. In fact, their vertical motion has evolved to be much more efficient than the horizontal propagation made possible by pseudopodia. Due to the laws of gravity, for the organism to be able to have a controlled vertical movement in the water it must have a significant difference in specific gravity compared to that of the ambient seawater. The mass of the shells and skeleton of the organism also must be considered in the mechanism by which the organism’s buoyancy is regulated. The Radiolaria’s heavy shells permit them to sink. In other words, gravitational forces could help explain how they manage their sinking behavior to maintain a certain sea level or be able to come back up closer to the surface. Planktons have evolved in their adaptations and techniques of buoyancy regulation through time. They’ve evolved to control the density of their body fluids with the use of an “ion pump”. The release of sulfate ions and retention of light ions such as chloride ions leads to a size reduction and increase in surface areas which allows for the delay of sinking. This is further aided by having large gas-filled vacuoles and high-buoyant lipid concentrations.
As for radiolarians, due to their heavy shells and skeletons, they secrete skeletal elements and frustules of opaline silica to achieve neutral buoyancy. When investigating buoyancy regulation of the organism, it is important to look at it as two components. First, the cells or protoplast altering in specific gravity. Secondly, the skeleton of a uniform specific gravity (Marszalek, 1981).
Specific gravity is defined by the ratio of the density of a specific substance to the density of a reference substance at a specific temperature and pressure.


If the ratio of density has a value bigger than 1 then it means that the specific substance is denser than the reference substance and if the ratio has a value less than 1 it signifies that the standard substance is denser than the substance in question (Torosian et al., 2016). Specific gravity is temperature dependent. Most substances will expand when temperature is increased and thus, have a decreased density (Stauffer et al., 2008). Hence, the specific gravity will decrease as well.
In this context, the specific substance is the cell or protoplast of Radiolaria, and the reference substance is water. A specific gravity less than 1 would result in the organism floating while a specific gravity bigger than 1 would lead to the organism sinking. In typical circumstances of static suspension, the combination of specific gravity of the heavy skeleton and the buoyant cell adds to around 1.026 which is approximately that of ambient ocean water (Marszalek, 1981).
When considering neutral buoyancy, specific gravity equilibrium must be achieved. In other words, the specific gravity of the protoplast must be equivalent to that of the seawater. Furthermore, negative and positive buoyancy can also be achieved. If the specific gravity of the protoplast is significantly smaller than that of the ocean water, positive buoyancy is reached. This could be a consequence of the organism secreting too many substances that are decreasing its mass. Thus, it will start moving in an upward vertical motion or floatation to the surface of the sea. Furthermore, negative buoyancy occurs when the specific gravity of the protoplast is significantly more than the specific gravity of the ambient seawater. Hence, sinking or downward movement is possible. However, the protoplast medium is primarily composed of water with very small quantities of proteins and other substances that are “dense”. Therefore, since it is practically impossible for the protoplast to have a significantly higher density than the density of seawater, negative buoyancy is rarely achieved. Hence, a cell component that has a very high specific gravity to seawater must be present in the cell to allow the organism to be negatively buoyant. Both negative and positive buoyancy are highly involved in the buoyancy regulation mechanism. That cell component is the mineralized shell and skeleton which has high density. In the case of Radiolaria, opaline silica is secreted from the cells to form spines and distinctive skeletons. The formation of heavy shells and skeletons allows for downward movement. For example, when the organism is positively buoyant, the mineralization of skeletons will be required to increase the specific gravity to that of the ambient seawater, which will allow it to attain buoyancy regulation. The mineralized skeleton and shells by opaline silica (Si(OH)4) were calculated to be around 2.0 (Marszalek, 1981).
In general, radiolarians contain cellular structures or compartments that hold fluid that is necessary to maintain hydrostatic pressure. Vertical movement of the hydrostatic organelles is effectuated partly through the formation and expulsion of CO2-saturated fluid. Specific gravity is temperature dependent. By consequence, so is buoyancy. As explained above, higher temperatures decrease the density of the organelles as well as the specific gravity. Conversely, lower temperature will do the opposite such that the cell or protoplast will be more compact, and denser, leading to an increase in specific gravity. Therefore, if the temporal changes are significant, it can cause positive or negative buoyancy.
Many species of Radiolaria have a complex skeleton structure that involves spines that could frequently reach around ten times the diameter of the shell of the organism. The spines are formed inside the organelle and then they precipitate in the near hours. Their main function is buoyancy regulation. They increase the resistance against forces exerted by seawater to keep the organism at the appropriate ocean depth. In other words, one could say that the spines play the role of a sea anchor for the Radiolaria. Furthermore, at times, the species that possess spines will shed their spines, secreting a carbonate crust as well. This crust will cover the shell of the organism. Thus, their heavy shells will increase in density resulting in sinking to greater water depths. This process happens before reproduction; gametogenic calcification is the name given to the process of the additional carbonate precipitation (Ohtsuka et al., 2015). Some plankton will sink to lower sea levels before releasing gametes to prevent “competition for survival and predation on gametes” that would happen closer to surface water level (Ohtsuka et al., 2015).
Radiolaria having a spherical shape is a recurring observation in different species. As Stoke’s law states, the rate of sinking is proportional to the square of the radius of spherical objects (Ohtsuka et al., 2015). Thus, an increase in the square of the radius leads to an increase in the sinking rate. Additionally, the size of Radiolaria affects buoyancy as well. Modeling its structure as a sphere lets us understand the effects of size on buoyancy. The surface area of the organism is directly proportional to its radius squared.

However, the volume is directly proportional to its radius cubed.

This means that as the Radiolaria get bigger, the volume will get larger much faster than the surface area. So, smaller Radiolaria will have a harder time lowering their density and thus their specific gravity than larger Radiolaria. This leads to larger Radiolaria being able to generate positive buoyancy much easier that smaller ones.
If the spherical shape is changed to a more planar or cylindrical shape, the sinking rate decreases. This could be observed in the way Radiolaria regulate their buoyancy through contraction and elongation to adjust the length of their pseudopodia. Hence, an elongation of pseudopodia results in the distortion of the Radiolaria’s spherical shape, which will delay sinking. Since the axopods and the reticulopodia greatly increase the surface area of the Radiolaria, they increase frictional drag forces. To add, when pseudopods are extended, they release air bubbles which contribute to the gas volume of the organism. Gas retention lowers the density of the organism which results in positive buoyancy and thus upward vertical motion. In turn, sinking can be aided by the retraction of their pseudopodia (Ryo et al., 2019). This occurs due to the sudden decrease in fluid drag experienced after retraction (Greco et al., 2023).
Distinctively, Acanthareans are observed to have a unique method of controlling buoyancy. This method involves a specialized structure called the periplasmatic cortex, which is a cytoplasmic membrane that can expand and contract rapidly. This function allows the periplasmatic cortex to change its volume, which in turn changes the volume of the cell (Suzuki & Not, 2015). Consequently, the cell’s specific gravity will change. So, the expansion of the periplasmatic cortex gives the Acantharia a positive buoyancy, allowing it to rise vertically in the water column. This structure works in tandem with two to twelve contractile myonemes, which are densely packed protein filaments that act similarly to animal muscles (Fig. 9). The periplasmatic cortex lies along the spicule (the spines of Radiolarian cells) in the same direction as the myonemes (Afanasieva & Amon, 2003). The myonemes work in contraction-relaxation cycles. They contract promptly to a fraction of their original length and then slowly relax back to being fully extended. The calcium concentration of the external medium plays a role in this contraction as well. Contraction occurs at 5×10-6 M of Ca and relaxation occurs at 10-7 M of Ca (Suzuki & Not, 2015).

Fig. 9 The black arrows identify groups of contractile myonemes which are attached to the periplasmic cortex and spicules of the Acantharia (Febvre-Chevalier & Febvre, 1994).
The control of buoyancy and thus vertical displacement is crucial for Radiolaria for numerous reasons. As mentioned above, some Radiolaria will shed their spines, secreting a carbonate, to sink lower in the ocean column before producing gametes to avoid a competition for survival. Additionally, Radiolaria live in symbiosis with algae, so they must spend daylight hours in the photic zone of the ocean at the optimal depth for algae to photosynthesize. Radiolaria have also been observed closer to the surface during calm seas and descend deeper during rough seas (Marszalek, 1982). So, vertical migration through buoyancy regulation is necessary for the daily life of a Radiolaria. The efficiency of these mechanisms has been improved and fine-tuned through hundreds of millions of years of evolution.
RADIOLARITE
Radiolarite is a type of chert-like sedimentary rock with a high silica content, formed by complex geological processes that offer a glimpse into ancient marine environments. Chert is a sedimentary rock that is primarily composed of microcrystalline quartz, which is the mineralized form of silica. Similarly, radiolarite is primarily composed of siliceous, preserved skeletal remains of radiolarians, making it a subtype of chert. A useful application for understanding the formation of radiolarite is for researchers to create transfer functions to reconstruct past ocean conditions, such as sea surface temperature and primary productivity (Chen et al., 2008).
Radiolarite Settlement
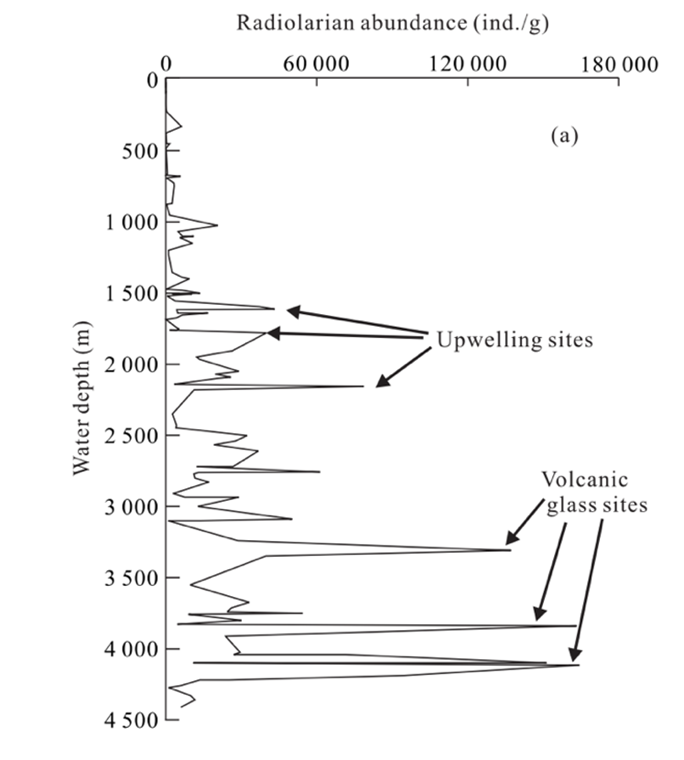
Once radiolarians die, the silica in radiolarian tests will begin to undergo dissolution as it descends to the ocean floor. According to Chen et al. (2008) (Fig. 10), radiolarians are well preserved in areas with a water depth of less than 4,200 m. In other words, this range is optimal for radiolarian fossilization. Beyond this depth (>4,200 m), evidence suggests greater dissolution in radiolarian shells.
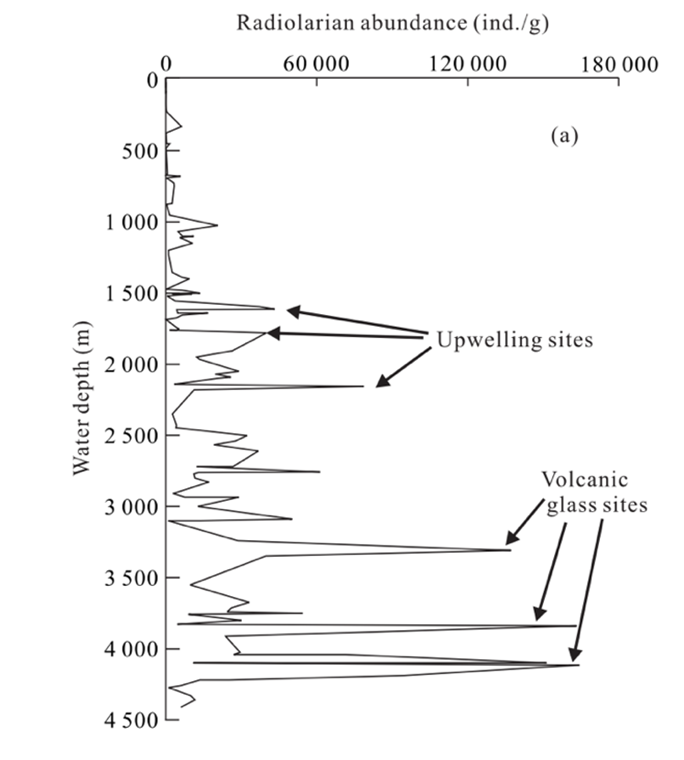
Fig. 10 Relations of radiolarian abundance (ind./g) with water depth in the South China Sea, noting peaks in areas where there is upwelling and volcanic activity (Chen et al., 2008).
It can take anywhere from 2 weeks to 14 months for radiolarians to descend water columns before reaching the ocean floor. Upon reaching the ocean floor, their tests begin to accumulate, forming layers upon layers of sediment, a process which can take many years (Takahashi, 1981). Numerous factors can determine their sedimentation rate. If radiolarians reproduce and die off in large numbers within a brief period, it can lead to a rapid accumulation of their tests. However, sedimentation is not just a function of biological factors; it is also determined by the physical properties of the tests themselves, determining their settling velocity (Vs) (Takahashi, 1981). According to Stokes’ law, the sinking speed (i.e., terminal velocity) of a spherical particle, for example, in radiolarian tests, in a Newtonian fluid is determined by the balance between drag forces and the force of gravity (Bridges & Robinson, 2020). Stokes’ drag (Fd), a frictional force, can be defined by the following equation:

μ(Pa · s) is the viscosity of the liquid, r (m) is the radius of the spherical object, and v (m/s)is the velocity of the particle (Bridges & Robinson, 2020).
By finding Stokes’ drag, the settling velocity can then be derived as a function of the density and size of radiolarian tests (Takahashi, 1983). According to Takahashi (1981), the density contrast between radiolarian tests and seawater is between 0.01g/cm3 and 0.05g/cm3. Correspondingly, the density can be used to determine the sinking velocity of the radiolarian, which Takahashi (1981) determined to be anywhere from 13 to 416 m/day. The sinking speed is defined by the following formula (6):
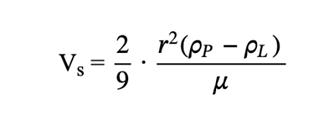
ρp (kg/m3) is the density of the particle, ρL (kg/m3) is the density of the fluid, r (m) is the radius of the particle, and μ (Pa · s) is the dynamic viscosity of the fluid (Bridges & Robinson, 2020).
Diagenesis
Over extended periods, the settling radiolarian tests will accumulate and undergo diagenesis, which is the process of the formation of radiolarite. Diagenesis involves key events such as compaction, recrystallization, and porosity reduction (Mat, 2023). As layers of sediment accumulate over time, the weight of the overlying sediments, known as overburden pressure, exerts force on the layers below. This pressure pushes sedimentary particles closer together, leading to a reduction in the void spaces between the layers, which reduces their porosity (Mat, 2023). Porosity is defined as the proportion of void space to the total volume of a rock or sediment. By decreasing porosity, it increases density and reduces permeability, which increases the strength of the rock (Hein & Obradović, 1989). Consequently, the pore-filling fluids are forced out, which can be described by Darcy’s law (Dandekar, 2013). Darcy’s law relates the flow of fluid through a porous medium to the porosity and permeability of the medium. Rocks with high porosity and permeability can act as reservoirs for fluids like water or hydrocarbons. Darcy’s equation can be defined as the following:
![\[q = -\frac{k}{\mu} \Delta p\]](https://bioengineering.hyperbook.mcgill.ca/wp-content/ql-cache/quicklatex.com-8de0c6e2e082f6ad5eebc8fe5dc029df_l3.png "Rendered by QuickLaTeX.com")
q (m/s) is the flux discharge per unit area, k (m2) is the permeability of the medium, Δp (Pa) is the pressure difference across the porous medium, μ (Pa · s) is the dynamic viscosity of the porous fluid (Dandekar, 2013).
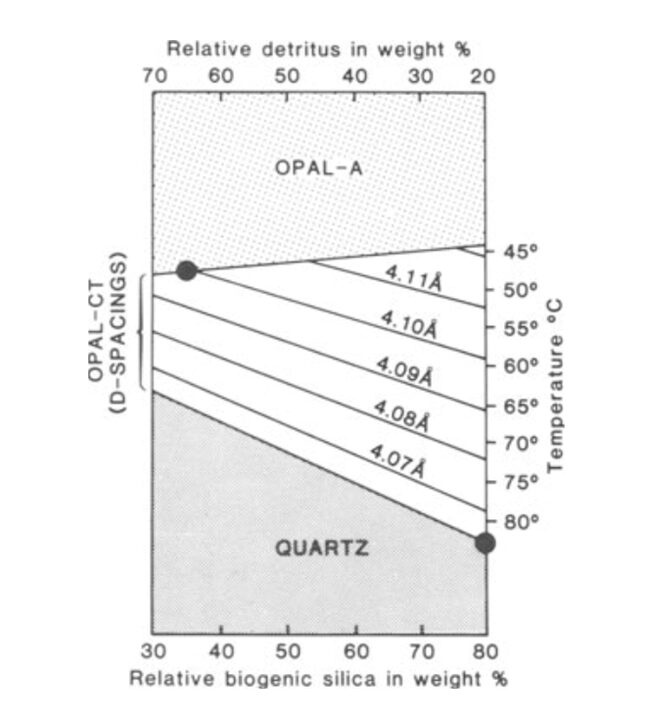
The silica from radiolarian tests can exist as various polymorphs. Factors such as temperature, pressure, and time determine the transformation of silica from one polymorph to another (Hein & Obradović, 1989). Initially, the sediment layers consist of opal-A, a biogenic opal that is a result of radiolarian tests (Haldar, 2020). It is amorphous, meaning it lacks a long-range ordered crystalline structure (Hein & Obradović, 1989). Opal-A is recrystallized and transformed into opal-CT (Cristobalite-Tridymite) via diagenesis with increasing burial depth at temperatures around 46 ± 2°C (Hein & Obradović, 1989). This phase consists of a mixture of disordered cristobalite and tridymite and is considered an intermediate between opal-A and quartz due to slight temperature increases at shallow depths (Haldar, 2020). With further prolonged burial, which continues to increase pressure and temperature to around 77 ± 5 °C, opal-CT can transform into chalcedony or even quartz. Both structures are more crystalline than the preceding forms of silica. Quartz, however, is the most stable form of silica at Earth’s surface conditions (Hein & Obradović, 1989; Haldar, 2020). As shown in Figure 11, with increasing temperature, diagenesis further progresses, transforming opal-A into quartz. Additionally, as the rock progresses through these crystalline structures, the d(101)-spacing (the distance between atomic planes in crystals) decreases, indicating the structure becomes more orderly and organized as diagenesis progresses (Hein & Obradović, 1989).
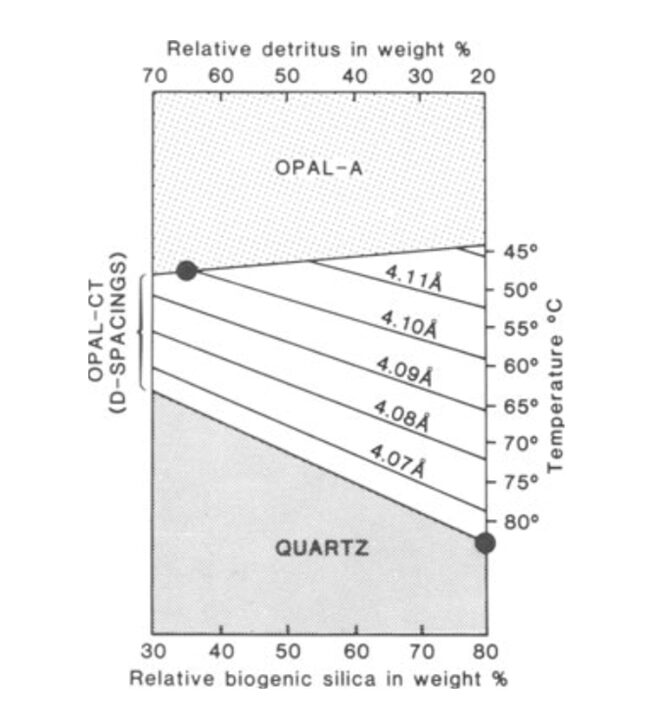
Fig. 11 Combined effects of temperature & rock composition during silica diagenesis (Hein & Obradović, 1989).
Through this mineral transformation, the texture of these sediment also evolves. Initially, the sediment is soft and porous, which is termed as radiolarian earth. As diagenesis progresses, the rock transforms into porcellanite and eventually into hard, dense chert (Hein & Obradović, 1989).
Tectonic Plate Activity’s Effects on Radiolarite Distribution
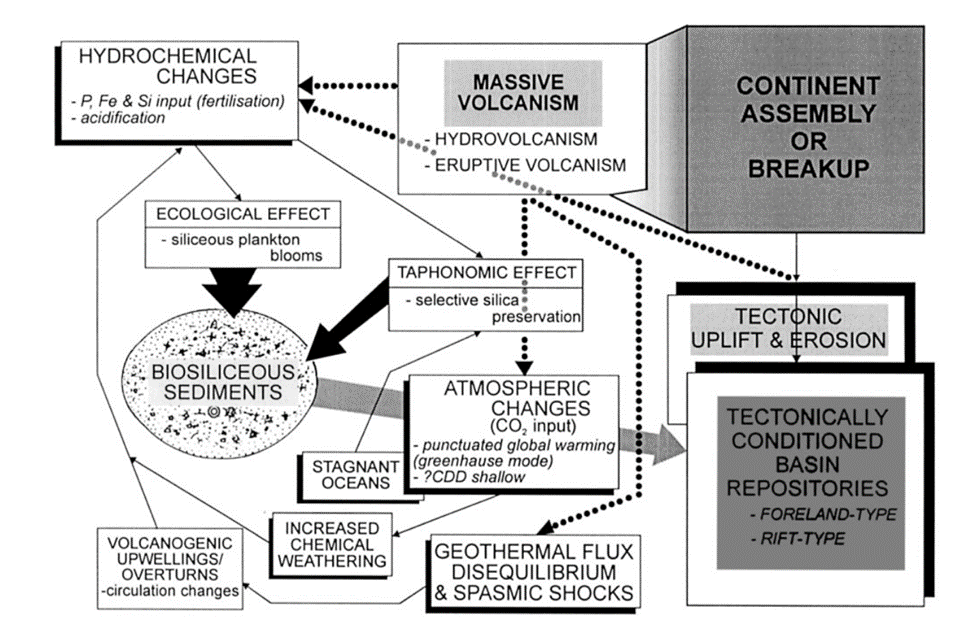
Radiolarites hold significance as sedimentary records, where their distribution can be used to infer past geological events based on the distribution of radiolarian microfossils. Their accumulation is usually favoured by geological conditions such as abyssal zones, volcanic eruptions, and upwelling activity, which are linked to tectonic activity (Chen et al., 2008). As noted, a crucial factor determining their distribution is tectonic movement, which can be visualized through a flow chart of events that promote biosiliceous sedimentation as shown in Figure 12. For example, tectonic processes can create closed basins, isolated areas of the ocean shielded from the usual influx of sediments from the land. Such environments are proposed to be ideal for the accumulation of biosiliceous-rich sediments in modern settings (Racki & Cordey, 2000).
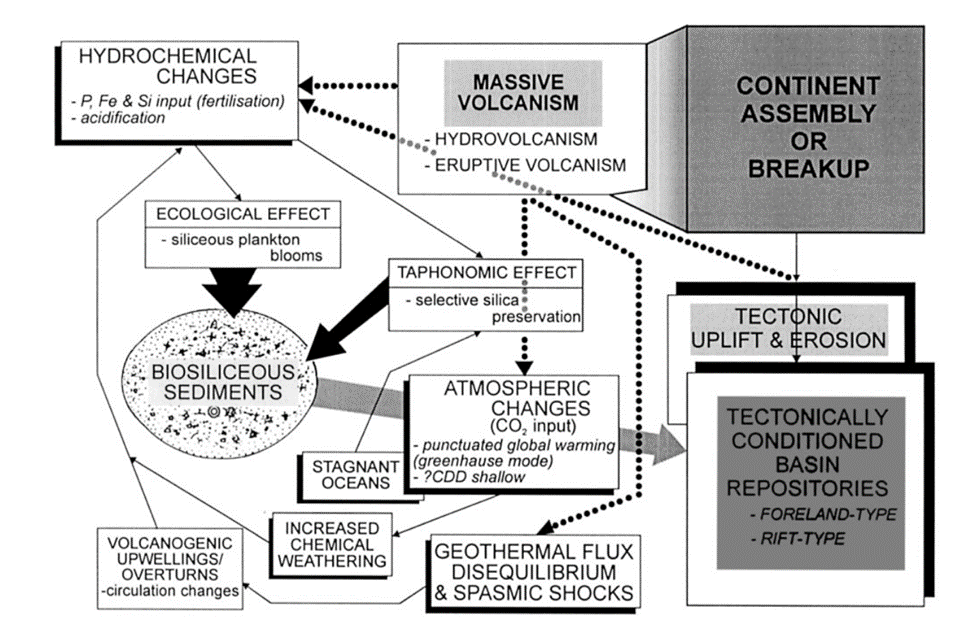
Fig. 12 Flow chart for the effects of volcanic and tectonic activity on biosiliceous sedimentation (Racki & Cordey, 2000).
Additionally, in a study conducted by Vecsei et al. (1989), they noticed that the cross-lamination observed in radiolarian chert at a microscopic level coincides with areas of the highest concentration of radiolarian tests. This provided evidence that the deposition of radiolarian ooze (Fig. 13) is influenced by the bottom-water current system, which causes selective radiolarian tests to settle out of water columns based on their hydraulic equivalence (Vecsei et al., 1989).

Fig. 13 Microscopic view of radiolarian ooze (Hata, 2011).
A key factor dictating bottom-water current is tectonic activity and the topological changes that occur with these movements, such as the opening or closing of oceanic gateways or changes in the size of ocean basins (Racki & Cordey, 2000). For instance, when landforms such as horst, an elevated block of earth’s crust, and graben, a valley like structure, are leveled they become like an elongated basin (Fig. 14), which occurs due to tectonic plate activities (Vecsei et al., 1989). Significantly, the change in topography allows bottom-water current and low-density turbidity currents to cover large areas, distributing radiolarian ooze. With this, Vecsei et al. (1989) was able to infer that during the time of the Oxfordian/Kimmeridgian boundary, increased tectonic activity caused changes to topography and the sedimentary environment, therefore showing that the distribution of radiolarians can be used to study and determine past geological events.
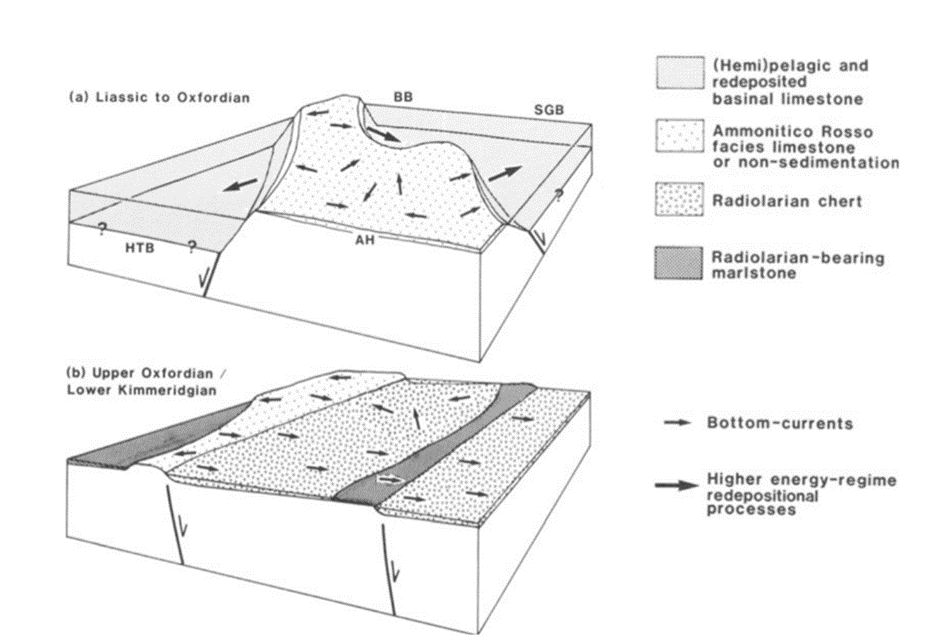
Fig. 14 Model for the evolution of underwater topography and sediment accumulation during the Jurassic period in the Northern Calcareous Alps. (a) During the early Liassic phase, there were elevated regions (horsts) and depressions (grabens). (b) By the transition between the Oxfordian and Kimmeridgian periods, this distinct topography transformed, moving from the horst and graben structure to a series of inclined blocks. As a result, most of the area became covered with Radiolarian chert, deposited by deep-sea currents (Vecsei et al., 1989).
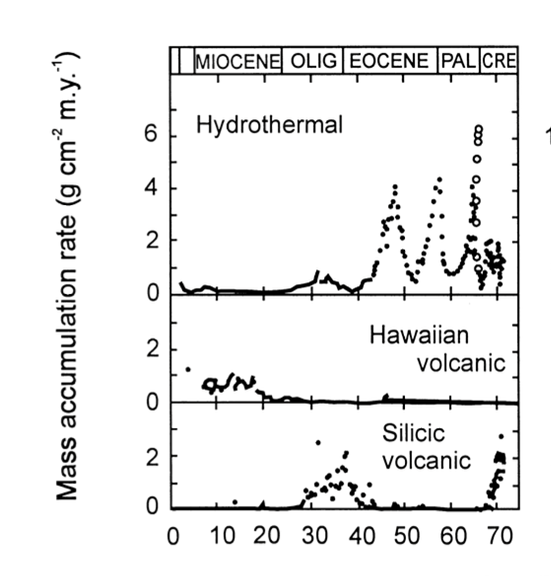
Volcanic activity also plays a vital role in radiolarite deposition. For instance, as shown previously in Figure 8, the peaks labeled upwelling sites and volcanic glass sites indicate that radiolarite formation is particularly favoured in volcanically active areas (Chen et al., 2008). Volcano-hydrothermal events refer to systems where volcanic activity drives the circulation of seawater near divergent plate boundaries, heating it up and causing it to pick up dissolved minerals before it is released into the ocean (D’Amico, 2019). Significant volcano-hydrothermal activities during plate-boundary reconfiguration are found to be hyper siliceous, indicating an increase in radiolarian accumulation in those areas. Furthermore, the silica content in hydrothermal fluids (Fig. 15) is much higher than in surrounding seawater, suggesting a higher abundance of radiolarian accumulation within those areas (Racki & Cordey, 2000).
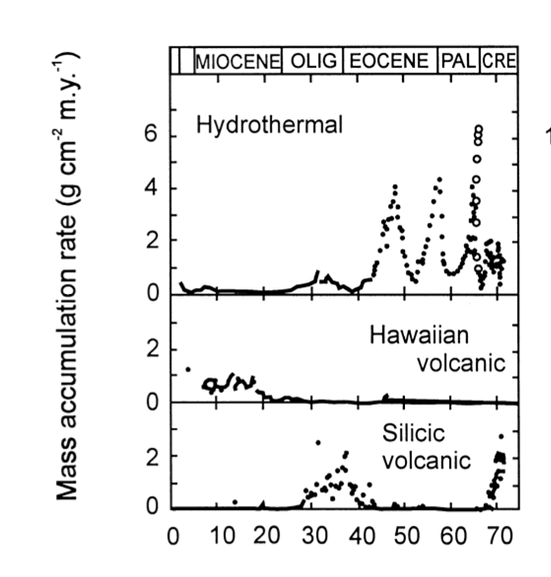
Fig. 15 The sedimentation records in different oceanic realms using geochemical markers. Notably, in the Eocene hydrothermal event, it increased siliceous productivity (Racki & Cordey, 2000).
CONCLUSION
Radiolaria are a widely diverse collection of unicellular eukaryotes. These zooplankton protists demonstrate a multitude of structurally elegant and physically complex processes in their oceanic environment. These various processes, systems, and structures have been explored in depth. The silica sponge spicules that compose Radiolaria skeletons can be viewed as biological fibers. The high refractive index of their spicules, in combination with a surrounding layer of cytoplasm with a lower refractive index, allows for light to be sent to the core more effectively, thus considering light absorption and scattering in water, which decreases the amount of available light to marine organisms. Since this process can be carried out in ambient temperature (whereas current commercial optical fibers must be made in high temperatures), it is of great inspiration to researchers in the field of optical fibres. Applications of this inspiration can be seen in development of optical fibre amplifiers and lasers. To add, this striking skeletal structure provides unique inspiration into lightweight yet stable structures that are of considerable interest to architects.
The locomotion of the organism, both horizontal and vertical, employ fundamental physics concepts. The use of axopodia as oars allows the organism to move through the water like a rowboat. In this motion, we can observe applications of laws of conservation of momentum, inertial forces and fluid drag forces. Although this forward propagation is not an efficient form of movement, their control of vertical motion is a rich example of efficiency triumphs that result from evolution. To be able to adjust their position in the water column, Radiolaria utilize a mosaic of intricate buoyancy regulation mechanisms. The use of ion pumping by way of substituting heavier ions for lighter ions allows them to lower their mass which in turn, also lowers their specific gravity to delay sinking. In addition, the organism can form heavy skeletons to generate negative buoyancy (sinking) and contrarily secret heavy components of its structure when positive buoyancy is needed. Gas retention by using vacuoles and pseudopodia extension are other major contributors to their ability to sink and rise in water as required. Evolutionary pressures that lead to the need to adjust their vertical position include their photosymbiotic relationship with algae, escape from rough sea conditions near the surface, and to ensure better probability of survival for their offsprings.
Formation of radiolarite at the bottom of the ocean provides geologists with a valuable fossil record. The settlement of Radiolaria depend primarily on the density and size of a given organism. The strength of these settlements is increased by a decrease in porosity and increase in density. These factors are, in turn, caused by an increase in pressure of the radiolarite caused by the accumulation of radiolarian tests. Rate of accumulation can be described by different laws of physics. Stoke’s law and Darcy’s law are two integral concepts that provide quantifications both of accumulation rate and the diagenesis process. Diagenesis can be described as the transformation of silica through various polymorphs that are guided by an increase in temperature and burial depth. Furthermore, a major effect that can be observed to alter distribution of radiolarian ooze (accumulation of radiolarian tests) is changes in topography caused by tectonic plate movements. So, geologists can study the distribution of radiolarite sedimentation to deduce an accurate record of ocean topography changes over time. Finally, these protists live in photosymbiosis with certain types of algae, meaning that they contribute to the ocean’s huge carbon sink. They are active contributors both while they live and when their tests settle at the bottom since they trap carbon dioxide with them. All things considered, radiolarians and their physical associations with the world provide indispensable insight into research and innovation across numerous fields of study.
REFERENCES
Afanasieva, M., & Amon, E. (2003). A New Classification of the Radiolaria. Paleontologica Journal, 37, 630-645. https://www.researchgate.net/publication/268485182_A_New_Classification_of_the_Radiolaria
Afanasieva, M. S., & Amon, E. O. (2014). Biomineralization of radiolarian skeletons. Paleontological Journal, 48(14), 1473-1486. https://doi.org/10.1134/S0031030114140020
Anderson, O. R. (1981). Radiolarian Fine Structure and Silica Deposition. In T. L. Simpson & B. E. Volcani (Eds.), Silicon and Siliceous Structures in Biological Systems (pp. 347-379). Springer New York. https://doi.org/10.1007/978-1-4612-5944-2_13
Atangana, A. (2018). Chapter 2 – Principle of Groundwater Flow. In A. Atangana (Ed.), Fractional Operators with Constant and Variable Order with Application to Geo-Hydrology (pp. 15-47). Academic Press. https://doi.org/https://doi.org/10.1016/B978-0-12-809670-3.00002-3
Biard, T. (2022). Diversity and ecology of Radiolaria in modern oceans. Environmental Microbiology, 24(5), 2179-2200. https://doi.org/10.1111/1462-2920.16004
Boltovskoy, D. (2017). Vertical distribution patterns of Radiolaria Polycystina (Protista) in the World Ocean: living ranges, isothermal submersion and settling shells. Journal of Plankton Research, 39(2), 330-349. https://doi.org/10.1093/plankt/fbx003
Bridges, S., & Robinson, L. (2020). Chapter 21 – Centrifuges. In S. Bridges & L. Robinson (Eds.), A Practical Handbook for Drilling Fluids Processing (pp. 475-488). Gulf Professional Publishing. https://doi.org/https://doi.org/10.1016/B978-0-12-821341-4.00021-X
Britannica, T. Editors of Encyclopaedia (2023). protist. Encyclopedia Britannica. https://www.britannica.com/science/protist
C, F.-C., & J, F. (1994). Buoyancy and swimming in marine planktonic protists. The Mechanics and Physiology of Animal Swimming, 13-26. https://doi.org/10.1017/cbo9780511983641.003
Chen, M.-T., Zhang, L., Xiang, R., & Lu, J. (2008). Radiolarian assemblages in surface sediments of the south china sea and their marine environmental correlations. Diqiu Kexue – Zhongguo Dizhi Daxue Xuebao/Earth Science – Journal of China University of Geosciences, 33, 775-782.
Chodniewicz, D., & Klemke, R. L. (2004). Guiding cell migration through directed extension and stabilization of pseudopodia. Experimental Cell Research, 301(1), 31-37. https://doi.org/https://doi.org/10.1016/j.yexcr.2004.08.006
Cooper, G. M., & Adams, K. W. (2023). The Cell: A Molecular Approach. Oxford University Press. https://books.google.ca/books?id=GNebEAAAQBAJ
Dandekar, A. Y. (2013). Petroleum Reservoir Rock and Fluid Properties. CRC Press. https://books.google.ca/books?id=tgTSBQAAQBAJ
Decelle, J., Colin, S., & Foster, R. (2015). Photosymbiosis in Marine Planktonic Protists. In (pp. 465-500). https://doi.org/10.1007/978-4-431-55130-0
Decelle, J., Probert, I., Bittner, L., Desdevises, Y., Colin, S., de Vargas, C., Galí, M., Simó, R., & Not, F. (2012). An original mode of symbiosis in open ocean plankton. Proceedings of the National Academy of Sciences, 109(44), 18000-18005. https://doi.org/doi:10.1073/pnas.1212303109
Ehrlich, H., Maldonado, M., Parker, A., Kulchin, Y., Schilling, J., Köhler, B., Skrzypczak, U., Simon, P., Reiswig, H., Tsurkan, M., Brunner, E., Voznesenskiy, S., Bezverbny, A., Golik, S., Nagorny, I., Vyalikh, D., Makarova, A., Molodtsov, S., Kummer, K., & Konstantin, T. (2016). Supercontinuum Generation in Naturally Occurring Glass Sponges Spicules. Advanced Optical Materials, 4. https://doi.org/10.1002/adom.201600454
Fenchel, T. (2013). Microorganisms (Microbes), Role of. In S. A. Levin (Ed.), Encyclopedia of Biodiversity (Second Edition) (pp. 299-308). Academic Press. https://doi.org/https://doi.org/10.1016/B978-0-12-384719-5.00094-0
Greco, M., Westgård, A., Sykes, F. E., Ezat, M. M., & Meilland, J. (2023). Uncovering hidden structures: previously undescribed pseudopodia and ectoplasmic structures in planktonic foraminifera. Journal of Plankton Research, 45(4), 652-660. https://doi.org/10.1093/plankt/fbad031
Haldar, S. K. (2020). Chapter 6 – Sedimentary rocks. In S. K. Haldar (Ed.), Introduction to Mineralogy and Petrology (Second Edition) (pp. 187-268). Elsevier. https://doi.org/https://doi.org/10.1016/B978-0-12-820585-3.00006-5
Hata, Y. (2011). SEM Photo of Radiolarian. In: Flickr.
Hein, J. R., & Obradović, J. (1989, 1989//). Siliceous Deposits of the Tethys and Pacific Regions. Siliceous Deposits of the Tethys and Pacific Regions, New York, NY.
Ichinohe, R., Shiino, Y., Kurihara, T., & Kishimoto, N. (2019). Active Floating with Buoyancy of Pseudopodia Versus Passive Floating by Hydrodynamic Drag Force: A Case Study of the Flat-Shaped Spumellarian Radiolarian Dictyocoryne. Paleontological Research, 23(4), 236-244, 239. https://doi.org/10.2517/2018PR023
Ishitani, Y., Febvre-Chevalier, C., & Febvre, J. (2016). Radiolaria. In (pp. 6). https://doi.org/10.1002/9780470015902.a0001985.pub2
Jungck, J. R., Wagner, R., van Loo, D., Grossman, B., Khiripet, N., Khiripet, J., Khantuwan, W., & Hagan, M. (2019). Art Forms in Nature: radiolaria from Haeckel and Blaschka to 3D nanotomography, quantitative image analysis, evolution, and contemporary art. Theory in Biosciences, 138(1), 159-187. https://doi.org/10.1007/s12064-019-00289-z
Lara, E., Dumack, K., García-Martín, J. M., Kudryavtsev, A., & Kosakyan, A. (2020). Amoeboid protist systematics: A report on the “Systematics of amoeboid protists” symposium at the VIIIth ECOP/ISOP meeting in Rome, 2019. European Journal of Protistology, 76, 125727. https://doi.org/https://doi.org/10.1016/j.ejop.2020.125727
Li, Z., Yin, D., Jiang, N., Wang, F., Ding, Y., & Li, F. (2022). Deformation and Failure Characteristics of Bimaterial Samples Consisting of Sandstone and Cemented Coal Gangue–Fly Ash Backfill under Uniaxial Loading. Minerals, 12(12), 1546. https://www.mdpi.com/2075-163X/12/12/1546
Marshall, W. F. (2020). Axopodia and the cellular “arms” race. Cytoskeleton (Hoboken, N.J.), 77(11), 483-484. https://doi.org/10.1002/cm.21642
Marszalek, D. S. (1982). The role of heavy skeletons in vertical movements of non‐motile zooplankton. Marine Behaviour and Physiology, 8(4), 295-303. https://doi.org/10.1080/10236248209387026
Mat, M. (2023). Diagenesis of Sedimentary Rocks. GeologyScience.com. Retrieved September 30th from https://geologyscience.com/geology-branches/sedimentology/diagenesis-of-sedimentary-rocks/?amp
Matul, A., & Mohan, R. (2017). Distribution of Polycystine Radiolarians in Bottom Surface Sediments and Its Relation to Summer Sea Temperature in the High-Latitude North Atlantic [Original Research]. Frontiers in Marine Science, 4. https://doi.org/10.3389/fmars.2017.00330
Nakamura, Y., Matsuoka, K., Imai, I., Ishii, K., Kuwata, A., Kawachi, M., Kimoto, K., Suzuki, N., Sano, M., Landeira, J., Miyamoto, H., Nishikawa, J., & Nishida, S. (2019). Current status on the taxonomy and ecology of plankton. Bulletin of the Plankton Society of Japan, 66, 22-40. https://doi.org/10.24763/bpsj.66.1_22
Pulman, C. (2005). The physics of rowing. University of Cambridge, 1-7.
Racki, G., & Cordey, F. (2000). Radiolarian palaeoecology and radiolarites: is the present the key to the past? Earth-Science Reviews, 52(1), 83-120. https://doi.org/https://doi.org/10.1016/S0012-8252(00)00024-6
Schowalter, T. D. (2016). Chapter 14 – Decomposition and Pedogenesis. In T. D. Schowalter (Ed.), Insect Ecology (Fourth Edition) (pp. 477-510). Academic Press. https://doi.org/https://doi.org/10.1016/B978-0-12-803033-2.00014-5
Sebastiano, D. A., Francesco, P., Salvatore, M., Roberto, I., Antonella, P., Giuseppe, L., Pauline, G., & Daniela, F. (2019). Chapter 12 – Ambient noise techniques to study near-surface in particular geological conditions: a brief review. In R. Persico, S. Piro, & N. Linford (Eds.), Innovation in Near-Surface Geophysics (pp. 419-460). Elsevier. https://doi.org/https://doi.org/10.1016/B978-0-12-812429-1.00012-X
Sharma, V., & Daneshian, J. (1998). Radiolaria as tracers of ocean–climate history. Current Science, 75(9), 893-897. http://www.jstor.org/stable/24101662
Sklar, L., & Dietrich, W. (2001). Sediment and rock strength control on river incision into bedrock. Geology, 29, 1087-1090. https://doi.org/10.1130/0091-7613(2001)029<1087:SARSCO>2.0.CO;2
Soden, M. (2011). The role of calcium in amoeboid movement: Results with fluorescent indicator dyes
Sundar, V. C., Yablon, A. D., Grazul, J. L., Ilan, M., & Aizenberg, J. (2003). Fibre-optical features of a glass sponge. Nature, 424(6951), 899-900. https://doi.org/10.1038/424899a
Suzuki, N., & Not, F. (2015). Biology and Ecology of Radiolaria. In S. Ohtsuka, T. Suzaki, T. Horiguchi, N. Suzuki, & F. Not (Eds.), Marine Protists: Diversity and Dynamics (pp. 179-222). Springer Japan. https://doi.org/10.1007/978-4-431-55130-0_8
Takahashi, K. (1981). Vertical flux, ecology and dissolution of radiolaria in tropical oceans : implications for the silica cycle.Woods Hole Open Access Server. https://doi.org/10.1575/1912/2420
Takahashi, K., & Honjo, S. (1983). Radiolarian skeletons: size, weight, sinking speed, and residence time in tropical pelagic oceans. Deep Sea Research Part A. Oceanographic Research Papers, 30(5), 543-568. https://doi.org/https://doi.org/10.1016/0198-0149(83)90088-2
Vecsei, A., Frisch, W., Pirzer, M., & Wetzel, A. (1989, 1989//). Origin and Tectonic Significance of Radiolarian Chert in the Austroalpine Rifted Continental Margin. Siliceous Deposits of the Tethys and Pacific Regions, New York, NY.
Vicker, M. G. (2000). Reaction–diffusion waves of actin filament polymerization/depolymerization in Dictyostelium pseudopodium extension and cell locomotion. Biophysical Chemistry, 84(2), 87-98. https://doi.org/https://doi.org/10.1016/S0301-4622(99)00146-5
Vicker, M. G. (2002). F-actin assembly in Dictyostelium cell locomotion and shape oscillations propagates as a self-organized reaction–diffusion wave. FEBS Letters, 510(1), 5-9. https://doi.org/https://doi.org/10.1016/S0014-5793(01)03207-0
Wagner, R. C., Jungck, J. R., & Van Loo, D. (2015). Sub-Micrometer X-ray Tomography of Radiolarians: Computer Modeling and Skeletonization. Microscopy Today, 23(5), 18-23. https://doi.org/10.1017/S1551929515000747
Yoshino, T., Matsuoka, A., Kurihara, T., Ishida, N., Kishimoto, N., Kimoto, K., & Matsuura, S. (2012). Application of Voronoi Tessellation of Spherical Surface to Geometrical Models of Skeleton Forms of Spherical Radiolaria. Zhou, X., Liang, P., Li, X., Guo, W., Zhang, X., & Yu, J. (2022). Filling Characteristics of Radiolarian Siliceous Shell Cavities at Wufeng-Longmaxi Shale in Sichuan Basin, Southwest China. Minerals, 12(12), 1545. https://www.mdpi.com/2075-163X/12/12/1545