


Introduction
The vertebrate brain has succeeded in the specialization of different traits and abilities through dividing it into two hemispheres, where the left hemisphere is better than the right one when conducting certain tasks, and vice versa. This specialization, called brain lateralization, also effects vision in vertebrates. In fact, visual lateralization, similarly to brain lateralization, defines the propensity of the eyes to specialize in their own respective functions (Manns and Ströckens, 2014). Additionally, it is believed to occur during the embryonic phase of an organism’s development (Manns and Ströckens, 2014). Hence, is theorized that this neuro-visual asymmetry is of genetic origin. However, many experiments and studies have shown that environmental factors can have significant impacts on the lateralization of an organism’s vision (Manns and Ströckens, 2014). Nonetheless, even though the concept of visual lateralization is common among vertebrates, it differs greatly from species to species. Natural selection has in fact permitted different vertebrates to utilize their lateralized vision in their own respective ways to assist them in their survival.
In this paper, we will be explaining the pertinence of visual lateralization in four different species: Pigeons, Chameleons, Dolphins, and finally, New Caledonian Crows.
Visual Lateralization in Pigeons
Visual Systems of Birds
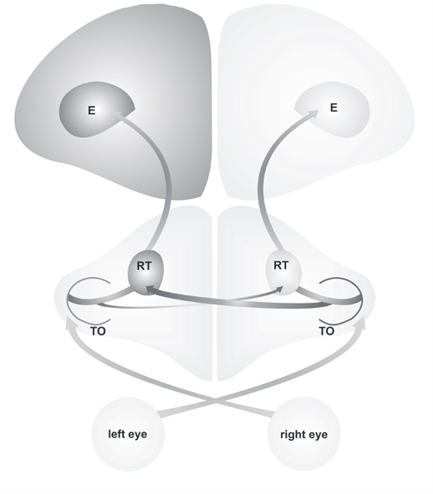
In birds, the left hemisphere of the brain is responsible for the analysis of little visual details, rule learning, and categorizing visual inputs that are received from the eyes. On the other hand, the right hemisphere is responsible for spatial attention and interactions with the environment (Manns and Ströckens, 2014). Experiments conducted on various bird species through the momentary covering of one eye have shown that the left eye is connected to the right hemisphere while the right eye is connected to the left hemisphere (Manns and Ströckens, 2014). This results in the crossing of the optic nerves, which is also referred to as the optic chiasm, and is actually a phenomenon observed in all vertebrate species (Bainbridge, 2009). Additionally, from what has currently been observed in the wild, the visual systems of birds and the way they are organized and lateralized can differ from one species to another.
These structural differences and visual pathway asymmetries are caused by differing amounts of light the right eye and the left eye receive during a bird’s embryonic development. This variation stems from the asymmetrical position of the embryo before hatching. In the egg, the bird embryo bends over to one side and turns its head right; this results in the right eye looking in the direction of the semitransparent eggshell. Contrarily, the left eye is obstructed by the body of the bird (Manns and Ströckens, 2014). This discrimination results in a significant variation in how much light each eye receives during embryonic development; light affects the right eye more than the left eye, resulting in both anatomical and functional lateralization in birds (Manns and Ströckens, 2014).
Visual Pathways of Pigeons
When discussing pigeons in particular, the tectofugal pathway, which is the visual pathway between the optic tectum and the forebrain that is responsible for orientation and attention, is lateralized, meaning there are left-right differences in the neural pathways (Reches and Gutfreund, 2009). In this case, this causes the left hemisphere of the pigeon’s brain to receive a better representation of information from the eyes, as can be seen in Fig. 1. However, it is worth noting that if an environmental condition obscures the embryo, depriving it from light, or if a genetic condition disrupts this sequence of events, the lateralization of the bird’s visual system will be affected as well; this can lead to some crucial asymmetries not being established (Manns and Ströckens, 2014). Hence, it is once again clear that light plays a crucial role in visual lateralization during the embryonic period.
Lateralization of Memory
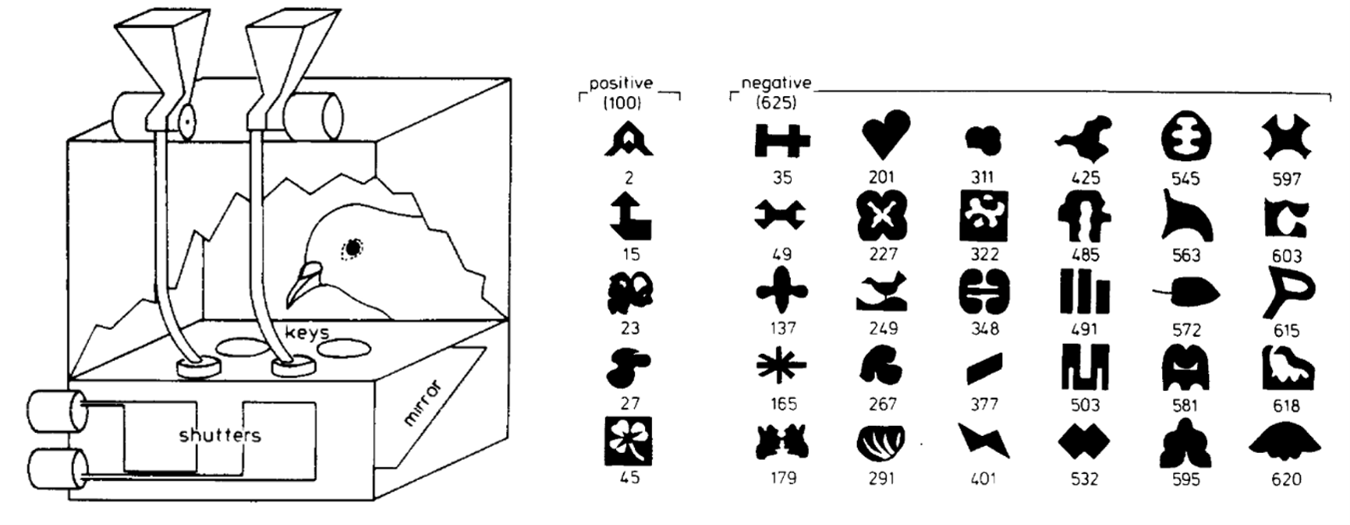
In pigeons, visual lateralization and its effects on behavior and the completion of various tasks was tested in an experiment conducted by Lorenzo von Fersen and Onur Güntürkün (2002). They built a system consisting of two automatic feeders placed above a cage, which could be activated by two buttons to deliver small amounts of food into the cage (Fig. 2). Each key was responsible for activating one feeder. A projector then displayed images of different shapes and patterns onto the keys. These shapes were grouped into two categories: 100 “positive” and 625 “negative” shapes (Fig. 2).

Pigeons were then placed in the cages and, in each trial, two random images were projected on the two buttons for 3 seconds (von Fersen and Güntürkün, 2002). The goal was for the pigeon to Fig. out and remember which shapes were positive and which were negative. If a pigeon pecked the button on which a positive image was projected, it would be rewarded with food; if both projected images were negative, the pigeon was expected to not peck and wait for the next round. If it incorrectly pecked, the cage went dark for 5 seconds. After 513 sessions, the pigeons had fully learned all positive and negative shapes. Then, the experiment was repeated while first covering the right eye and then the left eye. The results showed that pigeons correctly guessed the most when they used both eyes. However, when one eye was closed, they performed significantly better when their right eye was closed compared to when their left eye was shut (von Fersen and Güntürkün, 2002).
These aforementioned findings revealed a key organizational component of the optic neural pathway of pigeons; since the right eye is connected to the left hemisphere, these results show that visual lateralization affects memory functions in pigeons, as it causes differences in storage locations. The left hemisphere is the one responsible for visual memory processes, experimentally shown by the better performance of the pigeons when their right eye was open (von Fersen and Güntürkün, 2002). However, considering that pigeons still performed best when both eyes were open, it is clear that both hemispheres play a role in visual memory, despite the dominance of the left hemisphere. This shows that visual lateralization in pigeons does not cause some functions to completely be performed by the left hemisphere, but rather increase its role and function (von Fersen and Güntürkün, 2002).
Lateralization When Feeding
Further research, conducted by Güntürkün and others (2000), revealed the relation between the extent of visual asymmetry in the brain and how successful pigeons were in finding food (Güntürkün et al., 2000). In this experiment, 108 hungry pigeons were put through a grit-grain discrimination test, where 30 grains (food for pigeons) and 1000 grits are put in a container and the pigeon is given 30 seconds to find the grains. The performance of the pigeons was measured using both eyes, using only the left eye, and using only the right eye. The “absolute individual degree of lateralization” could be described using the asymmetry index (AI) (Güntürkün et al., 2000):
AI = right\space eye \space –\space left\space eye\space performance (\%)
(1)
The results showed once again that of pigeons performed best when utilizing both eyes. When comparing the results for one eye closed, pigeons with a higher AI, and consequently a larger degree of visual pathway asymmetries, performed better in differentiating grain from grit (Fig. 3). These results indicate that as visual asymmetry increases, the pigeon is more successful in finding food (Güntürkün et al., 2000). Furthermore, these results indicate that making one part of the brain specialize in object recognition possibly allows the computational speed of the whole process to increase (Güntürkün et al., 2000).

All in all, the structural left-hemispheric specialization of pigeons leads to differences in behavior and functions. Visual lateralization causes these birds to concentrate memory, pattern recognition, and object recognition in the left hemisphere of the brain, allowing for increased response accuracy.
Now, let us move onto our next case study on a very unique reptile!
Visual Lateralization in Chameleons
Chameleon Design Problem
Survival and reproduction are at the root of the circle of life in the animal kingdom. Every species has its own unique traits and skills and depends on their physical features to remain alive, by gathering food, and to reproduce. More interestingly, these survival skills are designed to compensate for the weaknesses of the species. With regards to chameleons, their main design problem is their extremely slow movement (Bradford, 2015). The latter puts them at risk in terms of predation and presents a big disadvantage for preying on their food (von Fersen and Güntürkün, 2002). To overcome this weakness, chameleons have developed amazing visual lateralization and incomparable eye features.
Uniqueness of Chameleon Eyes
When staring closely into a chameleon’s eyes, such as in Fig. 4, it is evident that they are formed differently from most other animals. In fact, these lizards lack a lower and upper eyelid, which is replaced by a cone-shaped single eyelid (Harris, n.d.). Chameleons, which are part of the family Chamaeleonidae, distinguish themselves from most of the animal kingdom by their ability to independently move their large-amplitude eyes (Lustig et al., 2012). In other words, a chameleon can be looking in one direction with its left eye, while looking in the completely opposite direction with the other.

Monocular Vision to Binocular Vision
Visual lateralization in the eyes of chameleons is used to capture food. Using both of their eyes independently (monocular vision), chameleons scan their surroundings in hopes of find prey (Harris, n.d.). Once this step is accomplished, a chameleon will rotate its head in the direction of the prey to be directly facing its target. This action allows them to use both of their eyes simultaneously by having them look in the same direction (binocular vision). This process allows for increase in accuracy; as mentioned by Rob Harris, “a chameleon uses binocular vision to increase his depth perception and aim to pinpoint accuracy as he shoots out his sticky tongue to catch the unsuspecting bug” (Harris, n.d.).
Interestingly enough, referring back to the physical aspects of eyes, chameleons possess a concave, also called negative, lens. As a consequence, these animals’ cornea is more powerful and allows them to focus precisely on the object they are looking at. This is how they can distinguish what they are looking at, a possible prey or something else (Miller, 2016).
360° Vision
While some animals have the ability to see in the dark, others possess different “superpowers”. As a matter of fact, it is almost impossible to take a chameleon by surprise because of their quasi-360° vision. As discussed earlier, this is made possible by their ability to independently move their eyes. More specifically, they have control over muscles that push and compress their eyes in and out and change it at different angles. Regarding these incredibly specialized muscles, “the eyeballs are mounted in twin conical turrets (like two upside down ice cream cones). Without a deep orbital socket to keep the eye from falling out (as in humans), the chameleon has evolved a thick, muscular lid. This lid surrounds each eye turret, leaving only the pupil exposed” (Miller, 2016). This mechanism provides a safe way for the eye to distance itself outwards from the conical turrets.
Extraocular Muscles
The chameleon is designed with 6 thin extraocular muscles, very much like all tetrapods. The presence of these muscles allows each chameleon eye to have a broader range of view; 180° horizontally and 90° vertically, as can be seen in Fig. 5 (Walls, 1942).
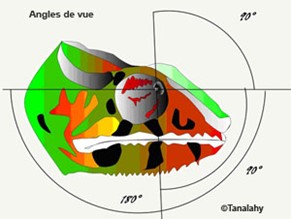
Furthermore, these muscles allow for a quicker response regarding the independent eye movement of these animals. As discussed earlier, the switch from the monocular to binocular view occurs due to an internal coupling in the neural substrate. “Further, neuronal binocular coupling was also observed during ocular accommodation in chameleons. Both eyes usually focus independently from each other. During prey fixation, however, accommodation is symmetrical in the two eyes even if one eye is covered” (Ott et al., 1998).
Now, onto our next case study on an animal that many could probably consider the most beloved aquatic species on Earth!
Visual Lateralization in Dolphins
Behavioral Uniqueness of Dolphins
Dolphins are particularly interesting species to consider with regards to their evolutionarily adapted visual lateralization. These animals participate in migration; an example is the Atlantic bottlenose dolphin, which travels north in warmer weathers and south in colder ones (Baxter, 2016). Consequently, these aquatic mammals are frequently exposed to a changing environment, and the lateralization of their visual pathway has adapted to work in favour of these circumstances. Additionally, as can be seen in Fig. 7, the specialization of the eyes in the completion of specific tasks works in favour of the dolphin’s monocular vision.

Have I Seen This Before?
Visual lateralization in dolphins is directly correlated to their ability to distinguish between new and familiar concepts and objects, which is crucial to their survival. A study was conducted by researchers Catherine Blois-Heulin and others in which they studied the response of dolphins to various stimuli, varying from very familiar objects, to mildly familiar objects, to completely unfamiliar objects. As can be seen in Fig. 8, what was observed was that these animals used their right eye to observe unknown objects, and their left eye to observe very familiar objects (Blois-Heulin et al., 2012). The researchers also observed that the time spent looking at a familiar object was significantly lower in comparison to the time spent looking at unknown objects. Undeniably, this difference is due to the effort the dolphin puts to try and attain some notion of familiarity from an object that does not strike it as familiar right away (Siniscalchi et al., 2012).

Considering that dolphins are vertebrates, their optic nerves cross just as they do in birds; the right eye connects to the left hemisphere of the brain while the left eye connects to the right hemisphere. Consequently, the way in which these mammals utilize their eyes differently at the execution of different tasks suggests that there exist important specializations in each hemisphere of the dolphin brain in relation to vision, as well the discrimination and analysis of what is being viewed. In fact, the results mentioned previously revealed that the right eye is the main contributor in the analysis of unfamiliar stimuli because the left hemisphere of the brain is involved in the detailed visual analysis of images and the completion of more intricate visuospatial tasks (Blois-Heulin et al., 2012). In terms of familiar objects, the left eye is preferred as detailed inspection of the visual stimuli is not required (Blois-Heulin et al., 2012).
A Social Species
Another study revolving around the visual laterality of social interactions in wild cetaceans revealed that bottlenose dolphins mostly use their right hemisphere, so their left eye, during flipper-to-body rubbing (F-B rubbing), which is when a dolphin rubs the body of another with its flipper (Sakai et al., 2006). After 382 video-recorded F-B rubbing episodes, the researchers noticed that this phenomenon was mostly conducted with the left flipper, and that it lasted longer than with the right flipper. Further analysis of this behavior suggested that the asymmetry in the F-B rubbing was caused by the laterality of the rubber, but also because both dolphins used the left eye during this activity, as it is an inquisitive behavior that does require analytic power (Sakai et al., 2006).
Migratory Behavior
Some researchers conducted further study concerning cerebral asymmetries in the visuospatial domain of two adult female bottlenose dolphins. The animals had to complete a task consisting of the following: they had to swim from a starting point in their tanks through a first hoop, then back to the starting point and through the second hoop, then back to the starting position, and finally through a third loop (Kilian et al., 2000). It was observed that the right eye performance of the animals was clearly superior to that of the left eye; this superiority is due to a left hemisphere dominance for the processing visuospatial information (Kilian et al., 2000).
Hence, visual lateralization in dolphins manifests itself in the treatment and analysis of stimuli. One such case is when it has to recognize whether an object is familiar to it; if it is, it is the left eye that responds, whereas if it is unknown, it is the right eye that responds. These experiments and studies pointed out some cognitive capacities of dolphins which are undeniably crucial for their survival given their migratory behavior. Also, the studies show the appearance of a preferred eye in the complete of various visuospatial tasks; the right eye is favored in responding to such stimuli that require visual localization (Kilian et al., 2000).
Last but not least, let us explore how a very intelligent bird species gives engineers a run for their money.
Visual Lateralization in New Caledonian Crows
Tool Use in the Animal Kingdom
Tool use is an exceptionally advantageous trait displayed by an array of members of the animal kingdom. From chimpanzees, to elephants, to octopuses, many species physically exemplify their high degree of cognitive ability by utilizing or crafting what is around them into objects that can serve them a specific purpose (Choi, 2009). We humans definitely are not the only engineers out there! One such an example of an animal that has fascinated scientists for its engineering abilities is the New Caledonian Crow (Hunt and Gray, 2004). Its propensity to manufacture tools, as well as its evolutionarily acquired visual characteristics that allow for effective tool use, will be discussed further in this section of our paper.
An Intelligent Species
Tool use in New Caledonian Crows has a direct impact on their survival, as this ability aids them in catching insects and larvae found in dead wood or vegetation, which are their primary source of food (Toscianko et al., 2012). These appliances can take on a variety of shapes and come from many different sources: New Caledonian Crows have been documented utilizing sticks and branches (Martinho III et al., 2014), leaf stems (Hunt and Rutledge, 2003), and even carefully cut hooked tools from the leaves of pandanus trees (Hunt and Gray, 2004) to dig and capture prey. The latter especially showcases the cognitive strength of these birds, as their pandanus tools have been shown to be more specialized and sophisticated than the tools manufactured by chimpanzees (Hunt, 2000). The method by which tools from pandanus leaves are crafted by New Caledonian Crows is showcased below in Fig. 9.
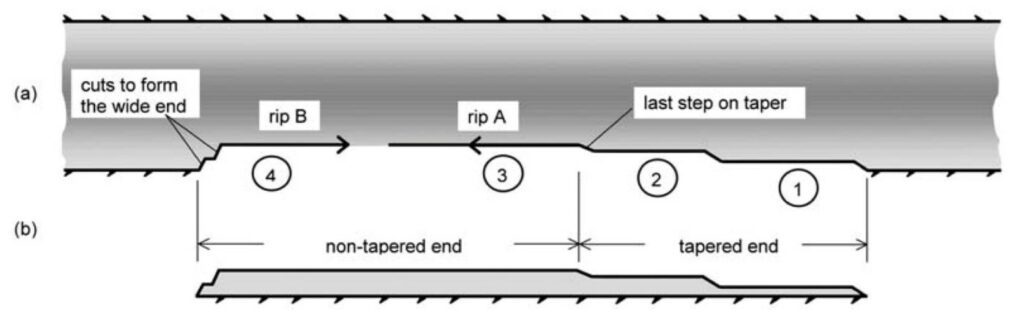
However, perhaps even more sophisticated than their manufacturing abilities is the way in which these birds use their tools. New Caledonian Crows hold their tools in their beaks and, most often, they have been observed holding their tools diagonally by positioning the non-working end of the instrument (the end that is not foraging for prey) against one side of their head, as can be seen in Fig. 10; in this way, the tool is properly secured and behaves like an extension of the beak (Rutledge and Hunt, 2004).
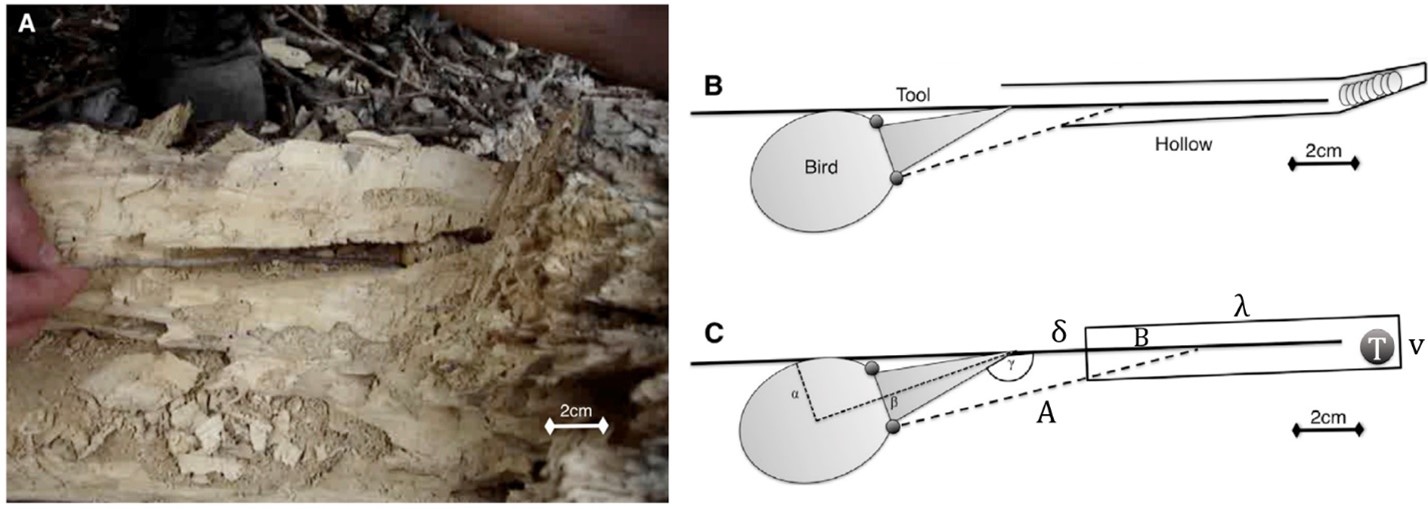
Additionally, it is worth noting that these birds do not press the non-working end of the tool against their eye; rather, they will position it below the eye located on the side of the head against which the tool is laying in order to maintain a clear picture of their surroundings and of the tool (Rutledge and Hunt, 2004). That being said, how does visual lateralization affect the New Caledonian Crow’s ability to use its tools? As we have discussed previously, evolution has led bird species to lateralize their vision, often giving preference in an individual for one eye over the other to complete certain tasks. New Caledonian Crows are no strangers to this, and in fact have evolved to utilize their lateralized vision during tool-conducted foraging. In order to explain this phenomenon, we will be repeatedly referencing Fig. 11, which explains the geometrical optics of New Caledonian Crow vision during foraging.

Lateralization of Tool Use
Firstly, it is worth explaining a few terms that will be reiterated throughout this explanation. The contralateral eye is the name given to the eye on the side of the head against which the non-working end of the tool is positioned. On the opposite side of the head is located what is called the ipsilateral eye (Troscianko et al., 2012). Evolution-induced visual lateralization has led New Caledonian Crows to possess one eye that is dominant, or preferred, over another (Martinho III et al., 2014). Additionally, research has shown that in terms of tool use, the side on which New Caledonian Crows will hold their tools can vary from individual to individual, but each of these individuals will consistently prefer to place their tools on the same side of the head throughout a lifetime. A comparable analogy is how primates can be right-handed or left-handed (Rutledge and Hunt, 2004). In fact, unlike the population wide lateralization observed in the three species mentioned above, the lateralization of tool use in New Caledonian Crows seems to vary from individual to individual (Rutledge and Hunt, 2004). The question now is how eye dominance affects the side on which a tool will be used, as well as whether it is the contralateral or ipsilateral eye that is dominant.
Consider Fig. 11c, showing a schematical depiction of a crow foraging for prey in a hole using some sort of stick. The contralateral eye may see directly into the hole, giving it a clear picture of the target prey (T) but also the very end of the tool. On the other hand, the ipsilateral eye’s potential field of vision (denoted by the dotted line labelled A), is directly dependent on various factors. The depth at which the ipsilateral eye may see the tool (B) will decrease if the distance between the tip of the beak and the opening of the hole (δ) decreases. Additionally, B will also decrease if the diameter of the hole (v) decreases, as well as if the depth of the target prey in the hole (λ) increases. As one can clearly see, the characteristics of the hole will have a significant impact on the ability of the ipsilateral eye to see the tool.
Can you now guess which eye is dominant in New Caledonian Crows? In order to always have a clear picture of their prey and their beak-held instrument, these birds will orient their dominant eye directly inside the hole and along the length of the stick. Thus, the dominant eye of New Caledonian Crows is the contralateral eye (Martinho III et al., 2014).
All in all, it is clear that these undeniably intelligent birds have evolved their tool-using abilities in collaboration with their lateralized vision; this has allowed visual lateralization to actually aid them in their survival by helping them see their prey and their tool with more ease when foraging.
Conclusion
To conclude, the brains of vertebrates have lateralized to give specializations to the left and right hemispheres, which, by consequence, has resulted in the lateralization of vision in many members of the animal kingdom. This process, called visual lateralization, has manifested itself in different species in various ways. Notably, visual lateralization has permitted the Pigeon, the Dolphin, and the New Caledonian Crow to specialize each eye in the performance of specific tasks, while the Chameleon has developed ways of independently using each eye to ultimately use them together and amplify its ability to perceive its prey. Thus, the eyes of an animal may look identical on the outside, but do not believe everything you see; the neural arrangement of these structures have allowed vertebrates to adapt to their surroundings and use their eyes in the most effective way possible for them. All in all, our three papers have showcased, from different angles, the incredible complexity of eyes; it is undeniable that engineers and scientists alike can truly learn from this incredible creation of nature.
References
Bainbridge, D. (2009). Beyond the Zonules of Zinn: A Fantastic Journey Through Your Brain: Harvard University Press. Retrieved from https://books.google.ca/books?id=cuCIyLmJkHoC
Baxter, R. (2016). Making a Move: Understanding the Dolphin Migration. Retrieved from http://www.biosphereonline.com/2016/05/03/understanding-dolphin-migration/
Blois-Heulin, C., Crével, M., Böye, M., & Lemasson, A. (2012). Visual laterality in dolphins: importance of the familiarity of stimuli. BMC Neuroscience, 13(1), 9. doi:10.1186/1471-2202-13-9
Bradford, A. (2015). Chameleon Facts. In. livescience.com. Retrieved from https://www.livescience.com/51061-chameleon.html
Choi, C. Q. (2009). 10 Animals That Use Tools. In. livescience.com. Retrieved from https://www.livescience.com/9761-10-animals-tools.html
Griffiths, S. (2015). Chameleons use their bulbous eyes to switch from stereo to mono vision and catch prey with pinpoint precision. Retrieved from https://www.dailymail.co.uk/sciencetech/article-3154881/Chameleons-use-bulbous-eyes-switch-stereo-mono-vision-catch-prey-pinpoint-precision.html
Güntürkün, O., Diekamp, B., Manns, M., Nottelmann, F., Prior, H., Schwarz, A., & Skiba, M. (2000). Asymmetry pays: visual lateralization improves discrimination success in pigeons. Current Biology, 10(17), 1079-1081. doi:10.1016/S0960-9822(00)00671-0
Harris, R. (n.d.). What Is Unusual About a Chameleon’s Eyes? In. animals.mom.com. Retrieved from https://animals.mom.com/unusual-chameleons-eyes-2288.html
Hunt, G. R. (2000). Human-like, population-level specialization in the manufacture of pandanus tools by New Caledonian crows Corvus moneduloides. Proceedings. Biological sciences, 267(1441), 403-413. doi:10.1098/rspb.2000.1015
Hunt, G. R., & Gray, R. D. (2004). Direct observations of pandanus-tool manufacture and use by a New Caledonian crow (Corvus moneduloides). Animal Cognition, 7(2), 114-120. doi:10.1007/s10071-003-0200-0
Katz, H. K., Lustig, A., Lev-Ari, T., Nov, Y., Rivlin, E., & Katzir, G. (2015). Eye movements in chameleons are not truly independent – evidence from simultaneous monocular tracking of two targets. Journal of Experimental Biology, 218(13), 2097-2105. doi:10.1242/jeb.113084
Kilian, A., von Fersen, L., & Güntürkün, O. (2000). Lateralization of visuospatial processing in the bottlenose dolphin (Tursiops truncatus). Behavioural Brain Research, 116(2), 211-215. doi:10.1016/S0166-4328(00)00273-4
Lustig, A., Ketter-Katz, H., & Katzir, G. (2012). Visually Guided Avoidance in the Chameleon (Chamaeleo chameleon): Response Patterns and Lateralization. PloS One, 7(6), e37875. doi:10.1371/journal.pone.0037875
Manns, M., & Ströckens, F. (2014). Functional and structural comparison of visual lateralization in birds – similar but still different. Frontiers in Psychology, 5(206). doi:10.3389/fpsyg.2014.00206
Martinho, A., Burns, Zackory T., von Bayern, Auguste M. P., & Kacelnik, A. (2014). Monocular Tool Control, Eye Dominance, and Laterality in New Caledonian Crows. Current Biology, 24(24), 2930-2934. doi:10.1016/j.cub.2014.10.035
Miller, A. (2016, 14 Sep 2016). Eyes Give 360° Vision. Retrieved from https://asknature.org/strategy/eyes-give-360-vision/
Ott, M., Schaeffel, F., & Kirmse, W. (1998). Binocular vision and accommodation in prey-catching chameleons. Journal of Comparative Physiology A, 182(3), 319-330. doi:10.1007/s003590050182
Pappas, S. (2012). Dolphins May Sleep-Talk in Whale-Song. In. livescience.com. Retrieved from https://www.livescience.com/18074-dolphins-sleep-talk-whale-song.html
Reches, A., & Gutfreund, Y. (2009). Auditory and multisensory responses in the tectofugal pathway of the barn owl. The Journal of neuroscience : the official journal of the Society for Neuroscience, 29(30), 9602-9613. doi:10.1523/JNEUROSCI.6117-08.2009
Rutledge, R., & Hunt, G. R. (2004). Lateralized tool use in wild New Caledonian crows. Animal Behaviour, 67(2), 327-332. doi:10.1016/j.anbehav.2003.07.002
Sakai, M., Hishii, T., Takeda, S., & Kohshima, S. (2006). Laterality of flipper rubbing behaviour in wild bottlenose dolphins (Tursiops aduncus): caused by asymmetry of eye use? Behavioural Brain Research, 170(2), 204-210. doi:10.1016/j.bbr.2006.02.018
Siniscalchi, M., Dimatteo, S., Pepe, A. M., Sasso, R., & Quaranta, A. (2012). Visual Lateralization in Wild Striped Dolphins (Stenella coeruleoalba) in Response to Stimuli with Different Degrees of Familiarity. PloS One, 7(1), e30001. doi:10.1371/journal.pone.0030001
Troscianko, J., von Bayern, A. M. P., Chappell, J., Rutz, C., & Martin, G. R. (2012). Extreme binocular vision and a straight bill facilitate tool use in New Caledonian crows. Nature Communications, 3(1), 1110. doi:10.1038/ncomms2111
von Fersen, L., & Güntürkün, O. (1990). Visual memory lateralization in pigeons. Neuropsychologia, 28(1), 1-7. doi:10.1016/0028-3932(90)90081-x
Walls, G. L. (2020). The Vertebrate Eye and Its Adaptive Radiation: Alpha Editions. Retrieved from https://books.google.ca/books?id=w8yBzQEACAAJ