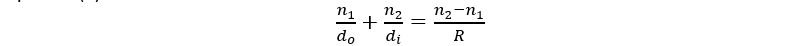
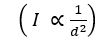
Abstract
This essay sheds light on a unique photoautotrophic prokaryote that presents a wide range of physical properties. Although billions of years old, cyanobacteria still ingeniously make use of their structural composition, mechanical processes, and optical properties to interact with their surroundings. With a strong gliding motility, a light sensing capacity, an ability to regulate buoyancy, and a potential formation of biofilm; cyanobacteria have it all. Each of these mechanisms is completely regulated by the cyanobacteria themselves and serves a specific purpose in enhancing their survival in constantly changing environments.
Introduction
Cyanobacteria, also known as blue-green algae in botanical terms (Whitton & Potts, 2012), are among the first inhabitants of the Earth (Mazard et al., 2016). This name originates from a blue color-inducing pigment called phycocyanin which most taxa produce (Whitton & Potts, 2012). Their history is debated, however, one estimate places cyanobacteria back 3500 Ma years (Whitton & Potts, 2002). They can vary immensely in habitat, ranging from freshwater and ocean habitats to dry environments. They can also survive under extreme conditions of heat, cold, or salinity. Cyanobacteria are prokaryotes that use photosynthetic pathways to create energy. Though it is impossible to truly know the exact number, there are an estimated 8,000 distinct species of cyanobacteria that inhabit the Earth. In 2022, only 5,185 species had been identified and classed (Mehdizadeh Allaf & Peerhossaini, 2022). Cyanobacteria are not just just varied, they are also incredibly abundant, with an estimated biomass of 3⋅1014 g⋅C (Whitton & Potts, 2012) and have been credited with the dramatic increase of oxygen in the Earth’s atmosphere 2.3 billion years ago (Mehdizadeh Allaf & Peerhossaini, 2022).
The cyanobacteria phylum encompasses both independent free-living bacteria and plastids. Plastids are the descendants of cyanobacteria that were incorporated into the ancestral lineage of the major photosynthetic eukaryotic group known as Archaeplastida. Subsequent endosymbiotic events led to the widespread distribution of plastids among various eukaryotic groups (Moore et al., 2019). Analysis of rDNA sequences in 2011 proposed that all current cyanobacteria may have a common unicellular ancestor. Molecular-based phylogenetic trees indicate that Gloeobacter violaceus, which possesses a light-harvesting mechanism limited to the outer cell membrane instead of internal thylakoids, is the closest living organism to this ancestral form. However, there is another hypothesis which suggests that the majority of contemporary cyanobacteria trace their lineage back to multicellular ancestors, with at least five instances of reversals to unicellular in their evolutionary history (Whitton & Potts, 2012).
Ranging in size between < 1 µm in diameter to 100 µm, cyanobacteria can also vary greatly in body composition and shape. There are unicellular, colonial, and multicellular structures (Figure 1). Unicellular cyanobacteria exhibit spherical, ovoid, or cylindrical cell shapes, and can form irregular or structured colonies held together by a mucous matrix (mucilage) produced during colony growth. Depending on the specific species, the number of cells within each colony can range from just a couple to several thousand. Following cell division in filamentous cyanobacteria, the individual cells stay connected, creating a linear structure referred to as trichomes. Depending on the order of cyanobacteria, the latter may or may not be enclosed by a mucous covering known as “filaments.” When trichomes fragment, they give rise to what are known as pseudobranches, a phenomenon observed across all cyanobacterial orders (Mehdizadeh Allaf & Peerhossaini, 2022).
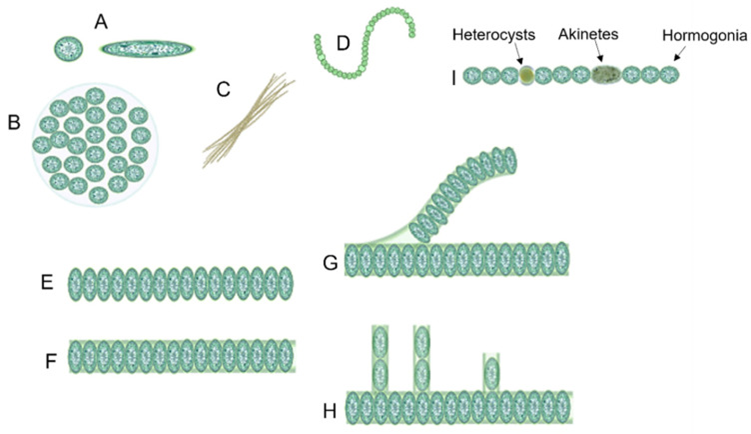
Figure 1. Structural differences between different forms of cyanobacteria: (A) unicellular (both spherical and ovoid), (B) colonial, (C) filamentous, (D) spiral, (E) unsheathed trichome, (F) sheathed trichome, (G) false branching, (H) true branching, (I) different cell types in filamentous cyanobacteria (Mehdizadeh Allaf & Peerhossaini, 2022).
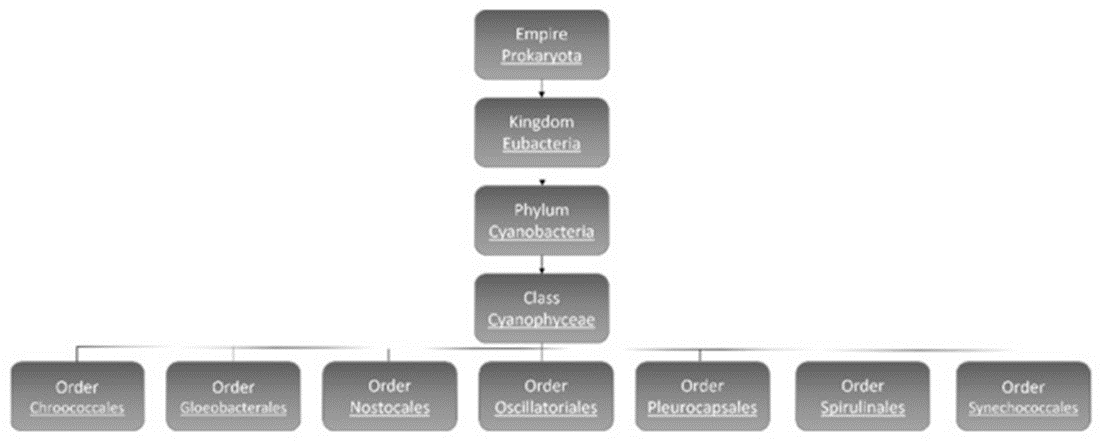
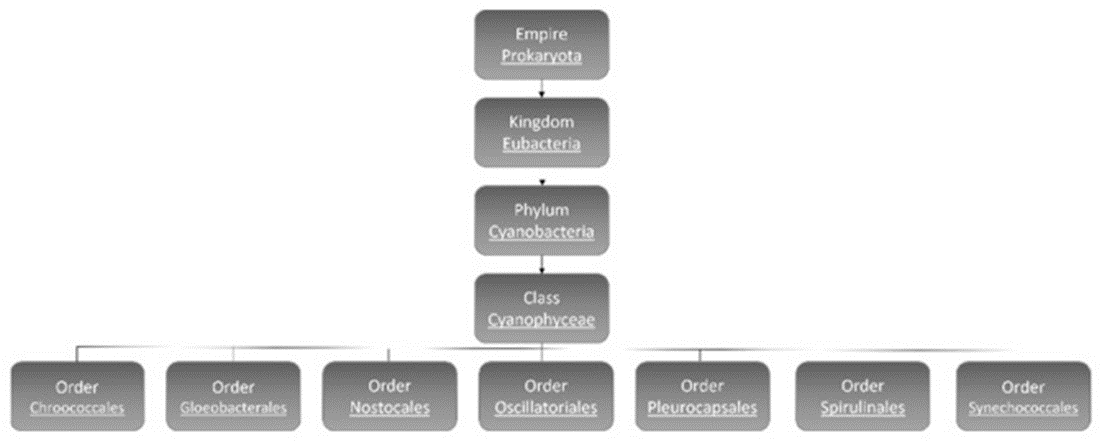
Cyanobacteria are subdivided into 7 orders: Chroococcales, Gloeobacterales, Nostocales, Oscillatoriales, Pleurocapsales, Spirulinales, and Synechococcales (see Figure 2) (Mehdizadeh Allaf & Peerhossaini, 2022).
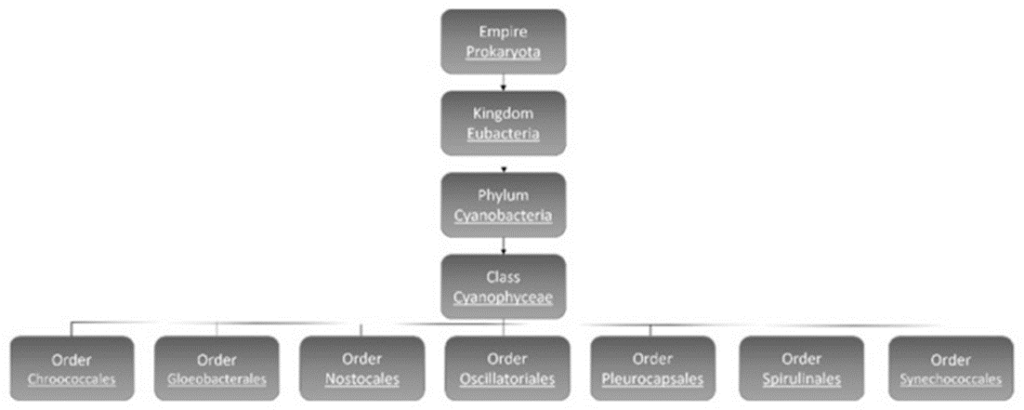
Figure 2. Classification of cyanobacteria (Mehdizadeh Allaf & Peerhossaini, 2022).
Though there is overlap between the characteristics of different orders of cyanobacteria, there are main distinctions between them. Order Nostocales are the filamentous type cyanobacteria, which include some that can fix nitrogen from the environment. Gloeobacterales are freshwater cyanobacteria, typically found either as single-celled organisms or in unstructured clusters, characterized by their rod-shaped morphology and the absence of thylakoids. Oscillatoriales and Pleurocapsales are the orders with the most benthic linear filamentous species with the latter having the possibility of being coccoid or semi-filamentous and can form complex colonies. Choococcales are unicelled coccoid and colonial, and they exist inside a gelatinous shell. Spiralinales are spiralized and filamentous and finally Synechococcales, the oldest group, consist of both unicellular and filamentous organisms (Mehdizadeh Allaf & Peerhossaini, 2022).
Cyanobacteria can have both positive and negative effects on their ecosystems. Among many other effects, notable ecological impacts are cyanobacterial blooms, which are large, aquatic surface colonies of cyanobacteria that can be very toxic to life below the surface by blocking sunlight, depleting oxygen content and interrupting the food chain (Havens, 2008). Cyanobacterial blooms increase in severity and frequency with increasing global temperatures due to climate change (Mehdizadeh Allaf & Peerhossaini, 2022). Cyanobacteria are an important part of terrestrial ecosystems where their presence in soil can increase the growth of flora because of their ability to fix nitrogen (Mazard et al., 2016).
Upon the long existence of cyanobacteria, there are some interesting mechanisms that have evolved in this organism. Cyanobacteria are photoautotrophs that can produce organic molecules from inorganic ones and generate energy through photosynthesis. They capture energy from light exposure, convert it into chemical energy, and store it as their own organic molecules (Svercel et al., 2011). This property makes sunlight one of the most important environmental factors for cyanobacteria’s survival. So, the cyanobacteria need to make sure the amount of sunlight they receive is enough to carry out photosynthesis but not enough to cause damage. How could such small organisms be able to do that? Cyanobacteria evolved to control the amount of sunlight they receive using a mechanism referred to as phototaxis that depends on their morphology and the environments they live in.
Phototaxis and Sensing Light
Spherical unicellular cyanobacteria can sense incident light as it passes through the cell and focuses an image on the opposite side of the cell (Schuergers et al., 2016). This phenomenon is illustrated in Figure 3 with bright spots corresponding to greater intensity of light. Cyanobacteria sense unidirectional light that is neither perfectly parallel nor perpendicular to the surface of the cell and move closer to the light source. This is a process called phototaxis, which is when an organism can respond to a light source by either moving closer or further away depending on which is more advantageous. In the case of two light sources facing opposite to one another, Syncechocystis sp. PCC 6803 moved towards the midpoint emphasizing their ability to adjust their behaviour while accurately detecting light and its intensity. The inverse square law (1) describes how the intensity of light diminishes as distance from the light source increases. This inversely proportional relationship highlights how cyanobacterial cells benefit from moving towards a light source if there is poor light intensity affecting the efficiency of photosynthesis.
I \propto \frac{1}{{d^2}}
(1) (Brownson, 2014)
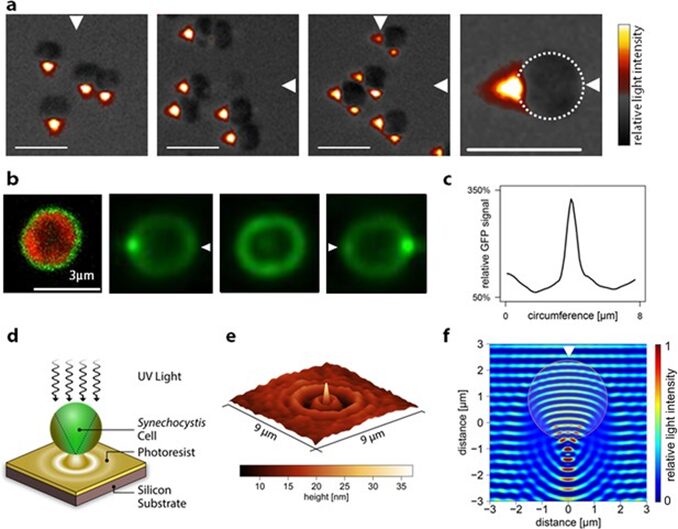
Figure 3. Images from the light incident on Synechocystis sp. PCC 6803 cyanobacteria demonstrate greater light intensity on the dorsal side of the cells. These cells function similarly to spherical micro-lenses by converging the incident light to produce an image on the opposite side of the cell (Schuergers et al., 2016).
The cell size is too small with a diameter of 3 µm to predict how the light will focus in the cell using typical ray diagrams (Schuergers et al., 2017) and therefore also the spherical interface equation (2).
\frac{n_1}{{d_o}} + \frac{n_2}{{d_i}} = \frac{n_2 - n_1}{{R}}
(2) (Ling et al., 2016)
Photonic nanojets are light beams characterized by narrow and high-intensity electromagnetic light where its beamwidth can be smaller than the optical diffraction limit (Heifetz et al., 2009). This provides greater resolution of the image of the light source than typical lenses are capable of. Photonic nanojets are formed in microspheres when its diameter is less than 4 µm. The phenomenon of photonic nanojets resembles what occurs in Syncechocystis sp. PCC 6803 cells due to their spherical shape and size, despite first appearing that they may act as a positive meniscus lens if the ventral side with incident light is isolated.
Filamentous cyanobacteria use their extended body length to analyze light gradients between multiple points and detect optimal locations (Biddanda et al., 2015). The filament shape also increases the surface area available for light exposure, adding to photosynthetic efficiency. Another tactic for increasing light optimization is interweaving many filamentous cells, creating clumps and mats. Having more cells increases the overall surface area of the light sensing “body” and increases the chances of detecting an optimal photic zone. These filamentous mats allow for both rapid horizontal and vertical locomotion which favours better responses to external stimuli.
A photophobic response is a protection mechanism that involves altering behaviour to minimize the impact of intense light. Cyanobacteria can perform phototaxis as a photophobic response to light that produces too great of an excitation from light absorption (Schuergers et al., 2016). When the intensity at the ventral side to incident light from a laser was greater than that on the dorsal side by a factor of 2-10, the Synechocystic altered its behaviour and exhibited negative phototaxis, meaning that it moved away from the light source. Cyanobacteria can sense light across the visible, ultraviolet, and infrared spectrum with photoreceptors (Wiltbank & Kehoe, 2019). These receptors are notably bile pigments and bile-binding photoreceptors, which can absorb specific wavelengths of light. Many photoreceptors are located strategically near effectors for signalling which send outputs based on the irradiance ratio between two wavelengths of light that are absorbed the greatest. Cyanobacteria can also increase the amount of light that is absorbed through chromatic acclimation. Light-harvesting antenna called phybilisome contain outer and inner rod segments. When there is a greater quantity of red light, the phybilisome phycocyanin concentration increases in the outer rod segment to absorb light of that wavelength range. When there is a greater quantity of green light, phytoerythrin, which is another type of phybilisome that has greater efficiency for its absorption, has a greater concentration in the outer rod.
Limiting Phototoxicity
Photoautotrophs like cyanobacteria must balance the dangers of phototoxicity and receiving enough light to produce energy from photosynthesis. CPh2 is a photoreceptor in cyanobacteria that regulates phototaxis according to the irradiance of specific wavelengths (Wiltbank & Kehoe, 2019). Positive phototaxis is inhibited by CPh2 with exposure to predominantly blue and low irradiant ultraviolet light. Cell aggregation is another tactic cyanobacteria employ to prevent phototoxicity by self-shading. For example, the cyanobacteriochrome SeSA promotes aggregation upon exposure to blue light. Some species of cyanobacteria also use the surrounding environment to protect from phototoxicity. The genus Oscillatoria cyanobacteria in springwater of the El Cajon Bay can seek shade from plants and other stationary objects (Biddanda et al., 2015). Oscillatoria and Microcoleus chthonoplastes can also vertically migrate upwards in a filamentous mat during the day and tend to be in more direct light. This contrasts with other species of cyanobacteria that move away from direct light sources.
In summary, the optical properties of cyanobacteria allow for precise light detection. By creating a sharp image of the light sources or increasing their surface area, cyanobacteria absorb the appropriate amount of light required for photosynthesis. However, there is still important information needed about how exactly cyanobacteria move to optimize their optical conditions.
Motility
The second part of phototaxis consists of the mechanisms that cyanobacteria use to move. The motility of cyanobacteria has been studied for over two centuries, and it has been observed that cyanobacteria use various mechanisms for movement, which can vary depending on the environment and species (Brahamsha & Bhaya, 2014). There are mechanisms used for moving on solid surfaces and mechanisms used for moving in liquid. On solid surfaces, the first pattern called gliding is observed in filamentous cyanobacteria and has the characteristic of moving at an average speed of about 2–4 μm per second without obvious change in morphology and occasionally reversing the direction of movement. The second pattern called twitching is observed in unicellular cyanobacteria which has the characteristic of moving at an average speed of about 0.05–1 μm per second with a jerky movement.
Motility on Solid Surface: Gliding
Organisms with the ability to glide do not have a locomotive organelle and do not display a noticeable change in morphology during movement. These two characteristics contribute to the mystery of gliding, as the fundamental mechanism behind it remains largely unexplained (Pate, 1988). A prominent model proposed to explain the driving force behind gliding suggests that it is caused by the secretion of mucilage by cyanobacteria.
Polysaccharide Secretion System
Filamentous cyanobacteria lack flagellum or other types of locomotive organelles. Instead, the propulsion of gliding is proposed to be generated by the secretion of polysaccharides. This mechanism is carried out by an organelle called a junctional pore complex (Hoiczyk & Baumeister, 1998). Cyanobacteria are gram-negative bacteria, characterized by a peptidoglycan cell wall located between their inner cytoplasmic membrane and outer membrane (Figure 4). The peptidoglycan layers of cyanobacteria range from 30 nm to 700 nm, which are considerably thicker than the 5 nm found in Escherichia coli. As a result, cyanobacterial cell walls create a significant permeability barrier, limiting the size of molecules that can pass through (Hoiczyk & Hansel, 2000). The junctional pore complex was also found within these peptidoglycan layers (Hoiczyk, 2000).
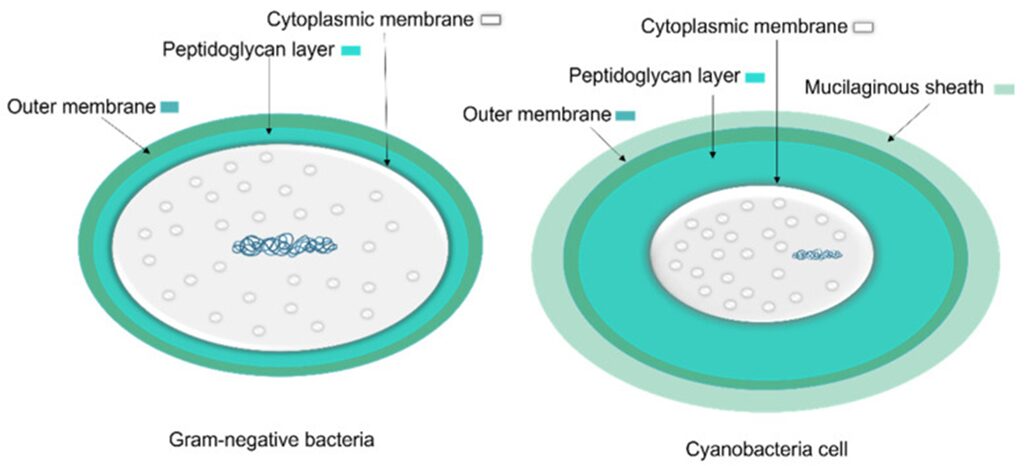
Figure 4. A comparison of general Gram-negative bacteria and cyanobacteria cell envelopes. The peptidoglycan layer in cyanobacteria is thicker than in other Gram-negative bacteria and the mucilaginous sheath is present in some taxa (Mehdizadeh Allaf & Peerhossaini, 2022).
Junctional pores (Figure 5) have diameters of 14-16 nm, are distributed in rows, and surround the filament on both sides of each cross wall or septum. The pores are attached to the tube-like part of the organelle which form channels that radiate outward from the cytoplasm, penetrate the peptidoglycan layer, and reach the septum at angles of about 30–40° relative to the plane of each septum. The total length of this structure is at least 70-80 nm, which is long enough to span the entire multilayered cell wall of the gliding filamentous cyanobacterium (Hoiczyk & Baumeister, 1998).
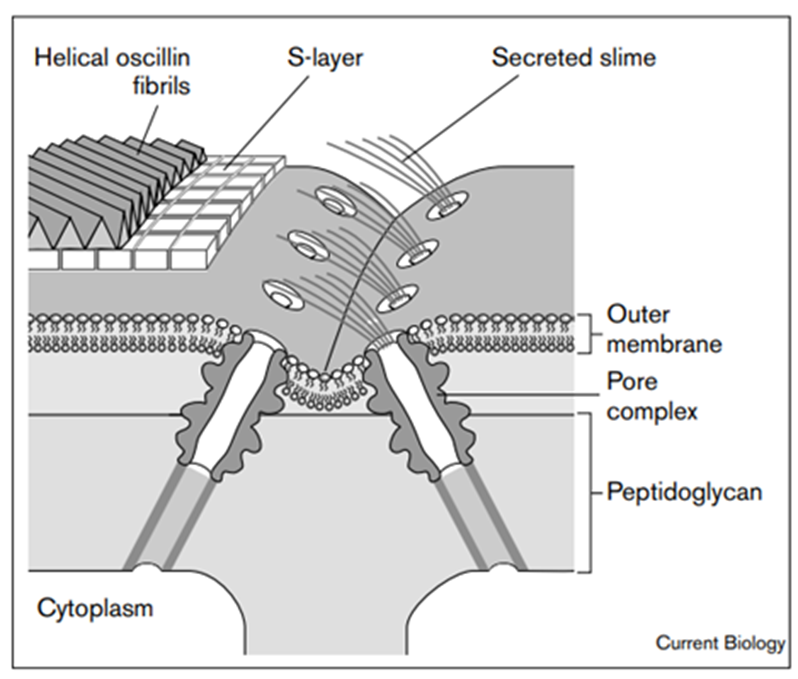
Figure 5. A model of the supramolecular architecture and activity of the junctional pore organelle in P. uncinatum. The pore organelle spans the entire cell wall complex (Hoiczyk & Baumeister, 1998).
Gliding by Rotating or Bending
The secretion of slime is proposed to generate the driving force for gliding in two ways. The first way is to cause the bacteria to helically rotate and move forward along its longitudinal direction (Figure 6). And the second way is to bend its long filamentous body toward the light source and move laterally (Figure 7). The different results of moving pattern generated by slime secretion is caused by the structural differences between the species of cyanobacteria.
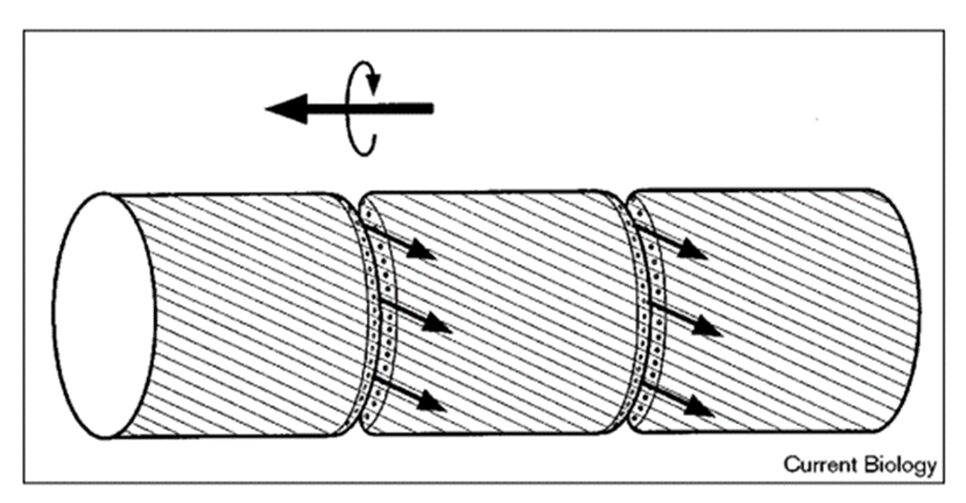
Figure 6. Schematic illustration of the ‘surface-mucilage’ hypothesis for gliding motility in cyanobacteria. The pore organelle sets (indicated by small circles at the cross walls of the cells) steadily secrete mucilage (indicated by small arrows) that flows in tight contact with the filament surface and adheres to the substrate surface. Secretion and flow of the mucilage causes translocation of the filament, and the helically arranged surface fibrils formed by the protein oscillin guide the rotation of the filament. The frequent reversals of movement may result from the change in the direction of mucilage flow, caused by the alternation of the set of pores used (Hoiczyk & Baumeister 1998).
For the rotating motion, take the genus Phormidium cyanobacteria as an example. The researcher hypothesizes that the rotation is caused by the fibrils that are arranged helically on the surface of the Phormidium bacterium (Figure 6). After mucilage is secreted from the junctional pore complex, it attaches to the surface of the filament and adheres to the substrate surface. Since there are fibrils arranged helically on the surface of the filament, the flow of mucilage is also directed helically. This helical flow of mucilage causes the filament to rotate and glide (Hoiczyk & Baumeister, 1998).
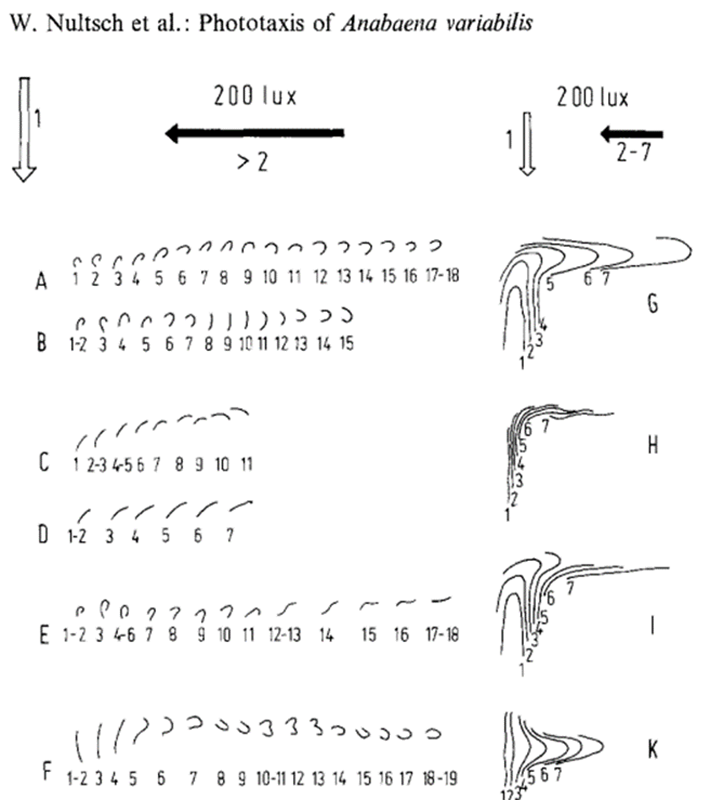
Figure 7. Change of movement direction of the genus Anabaena cyanobacteria trichomes at 2001x. Light arrow: initial light direction; dark arrow.” new light direction. 1-19 denote the positions of the trichome, in 1 min intervals after changing the light direction. A, B, show the behaviour of U formed trichomes. C, D show the behaviour of straight trichomes. E, U, formed trichome becoming straight after the change of light direction. F, straight trichome becoming U formed after the change of light direction.
For the bending motion, take Anabaena as an example. The surface of Anabaena is not covered by the helical fibrils, so the secretion of mucilage would not cause it to rotate. The arrangement of its junctional pores in girdles and its ability to secrete slime from one flank together allow the filament to bend and move laterally (Hoiczyk & Baumeister 1998).
Twitching
Unicellular cyanobacteria have a different morphology from filamentous cyanobacteria in that that they use organelles called pili to move (Figure 8) (Maier & Wong, 2015).
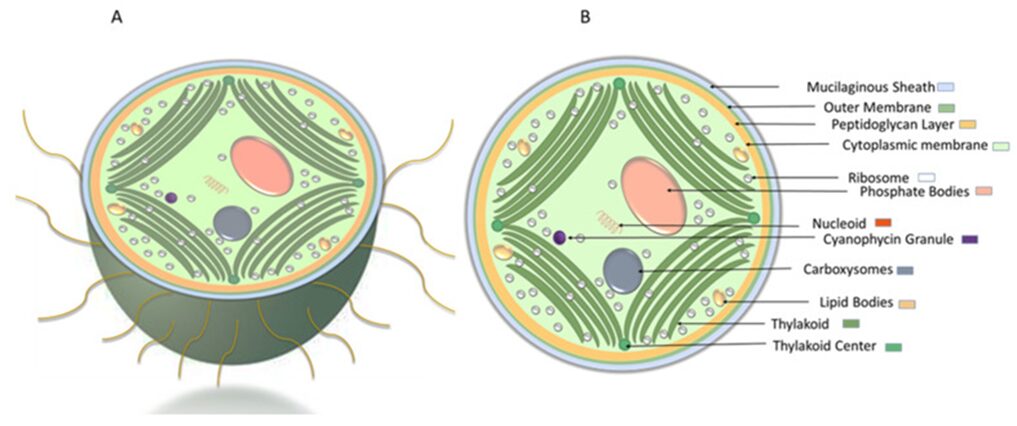
Figure 8. Schematic representation of Synechocystis cell morphology. (A) cross-sectional view of Synechocystis cell with pili. (B) ultrastructure of Synechocystis cell (Mehdizadeh Allaf and Peerhossaini 2022).
Type IV pili extend several micrometers from their body and can elongate via polymerization and retract via depolymerization. The movement of unicellular cyanobacteria is driven by the retraction of type IV pili after they adhere to an object (Figure 9) (Maier & Wong, 2015). A single type IV pilus can generate a retraction force exceeding 100 pN, making it one of the strongest molecular machines characterized to date. In organisms like N. gonorrhoeae (gonococcus) and M. xanthus, this force can be amplified tenfold by forming a bundle with 8 to 10 pili (Biais et al., 2008).
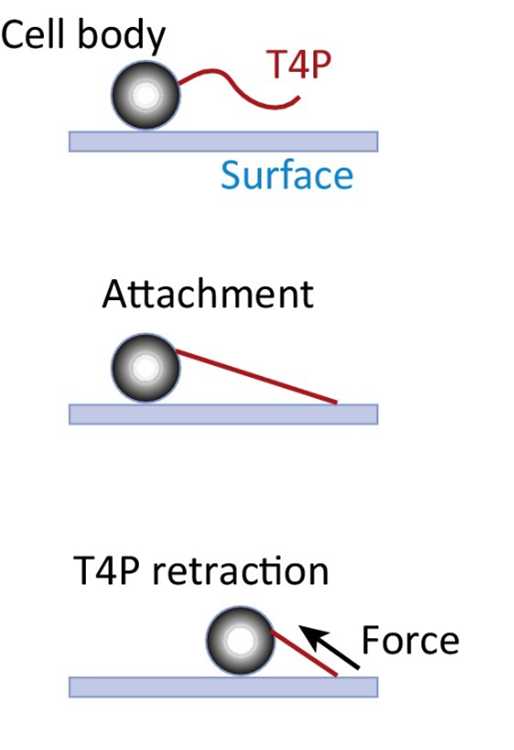
Figure 9. The type IV Pilus (T4P) Machine Mediates Twitching Motility. (A) By cycles of T4P polymerization, surface adhesion, and retraction, the T4P pulls the cell body along surfaces (Maier & Wong, 2015).
Although the set of proteins required for motility and the molecular mechanism of force generation are well conserved through unicellular cyanobacteria species, the differences in habitat and phylogeny determine the distinct mechanisms that they use to coordinate multiple pili (Maier & Wong, 2015).
The genus Neisseria cyanobacteria exhibit a mechanism in which type IV pili are generated randomly on their cell contou. The direction of each movement is the result of the net force caused by the retraction of pili from several directions (Figure 10). This mechanism makes the moving pattern of Neisseria species to be a random walk with directional persistence at a short time scale. If the density of pili is too high in one cell (≈30 pili per cell), the cell is likely to be trapped frequently because the forces are more likely to be balanced (Maier & Wong, 2015).
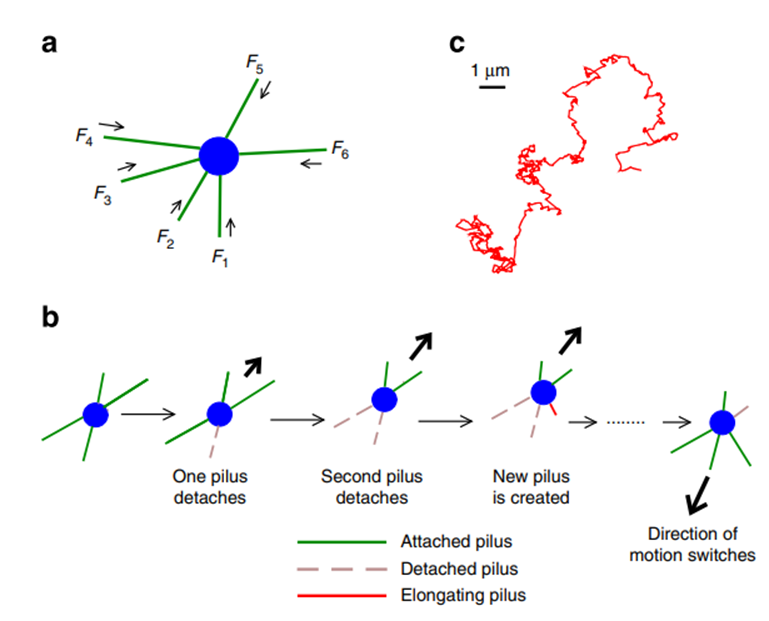
Figure 10. Two-dimensional tug-of-war mechanism for twitching motility: (a) A bacterium with pili emanating in different directions. The arrows indicate the retraction of pili; the forces experienced by the pili are denoted by Fi and taken as positive if they oppose the direction of retraction. (b) Basic tug-of-war mechanism: an imbalance in pulling forces in different directions is amplified by detachment of those pili that experience large force, resulting in directed motion. The direction of motion can change stochastically. (c) A typical trajectory as obtained from the simulations for the case of random movement (Marathe et al., 2014).
Another species, P. aeruginosa has a rod shape and generates type IV pili at the cell poles (Maier & Wong, 2015). This mechanism differs in that it can switch between two different surface motility mechanisms. In the ‘crawling’ mechanism, bacteria move lengthwise, parallel to the surface with high directional persistence. The other mechanism, ‘walking’, is a movement directed upright, perpendicular to the surface, by which the bacterium moves with low directional persistence. Walking bacteria also have a shorter persistence length (Lp) (2 μm) than crawling bacteria (6 μm). As a result, although walking bacteria have a higher instantaneous velocity [mean 71 ± 2 nm/s (SEM)] than crawling bacteria do (2 nm/s), crawling bacteria move further on long time scales because of the Lp. Crawling enables directional motion; walking enables rapid local exploration. These trends were preserved in wild-type bacteria (Gibiansky et al., 2010).
Overall, cyanobacteria move on surface by gliding or twitching. The lack of obvious locomotive organelle for the species of cyanobacteria using gliding has led to researchers hypothesizing that the secretion of slime drives this kind of motion by either causing the organism to rotate or bending its body and move laterally. The basic mechanism of twitching conserved through species of cyanobacteria, which is a cycle of elongation, attachment, and retraction of the type IV pili on its body. However, the way to control the direction of the whole movement varies from species to species. Based on the distribution of their type IV pili, the direction of the movement could be the result of the net force generated by all the retracting type IV pili at a given point in time or having two or more moving modes that either have a different preferring direction to move and switch between them.
The importance of buoyancy regulation in aquatic environments
For mobility in aquatic environments, cyanobacteria, like the rest of the phytoplankton community, rely on vertical migrations. To adjust the intensity of the light they receive, cyanobacteria control their depth in water through buoyancy regulation. Buoyancy is the tendency of an object to float when immersed in a fluid. The pressure applied by a fluid increases with depth. This means that the pressure at the bottom of an object is higher than the pressure at the top. This difference in pressure results in an upward buoyant force exerted by the fluid on the object which opposes the downward force exerted by the presence of gravity. For the cyanobacteria completely submerged in water, the buoyant force formula simplifies to the following equation (3) where FB represents the buoyant force and depends on V the volume of the organism, r the density of the fluid (here water), and g the gravitational acceleration.
F_B = V \rho g
(3) (Cochran & Heinrich, 2021)
The buoyancy attribute of cyanobacteria is provided by a specific structural element: gas vesicles. A gas vesicle is a hollow tube-shaped cylindrical structure closed by hollow conical caps at each end and it contains a gas-filled space. It is made of ribs of proteins perpendicular to the long axis. To ensure gas permeability, gas vesicles are not inflated but self-assembled from biconical initials. Thanks to their hydrophobic inner surface, these structures are also impermeable to water since, even when completely immersed in aquatic environments, the gas vesicles of cyanobacteria are never filled with water (Reynolds, Oliver, & Walsby, 1987). All cyanobacteria species share a common gas vesicle shape that only differs in size. The diameter of the cylinder is usually between 45 nm and 110 nm. Gas vacuoles, the aggregates of gas vesicles, are visible under a microscope, but the gas vesicles themselves are too small (Walsby, 1992). Figure 11 shows a clear image of cyanobacterial gas vesicles obtained by electron microscopy.

Figure 11. Electron micrographs of gas vesicles from Aphanizomenon flos-aquae, a species of cyanobacteria (Daviso et al., 2013).
Mechanical properties of gas vesicles
Unlike gas bubbles, the gas vesicles of cyanobacteria are rigid structures characterized by an elastic compressibility. This property refers to the change of an object’s volume when pressure is applied on it.
To have a better understanding of the stresses in the wall of gas vesicles and how much they can be compressed or deformed, we may refer to Young’s modulus through the simplified Newman & Searle equation represented below (4). On the left-hand side, dV/V represents the volume change. It is a resultant of the pressure P applied on the right-hand side of the equation. It is also dependent of the cross-sectional radius R of the cylinder, the wall thickness h, and Poisson’s ratio σ. Poisson’s ratio is defined as the ratio of the change in width per unit width of a material, to the change in length per unit length of the same material due to the strain of the stretching force (Ebnesajjad & Khaladkar, 2005). When all the parameters are known, determining the elastic modulus Y of the gas vesicle wall protein becomes relatively simple. However, we should note that this method is not entirely accurate. This calculation assumes that the cylindrical wall is homogeneous, of even thickness, and with flat ends which is not applicable to gas vesicles (Walsby, 1982).
\frac{dV}{V} = \frac{PR(4\sigma - 5)}{2Yh}(4) (Walsby, 1982)
The elastic compressibility of gas vesicles indicates that the structure sees a decrease in its volume when low pressures are applied. Conversely, under higher pressures, gas vesicles irreversibly collapse. The “critical” pressure is the minimal difference of pressure between the interior and exterior of the gas vesicle that causes the structure to collapse (Walsby, 1994). The elastic modulus Y of a gas vesicle is a measurement of its low compressibility which is very important in natural habitats. The hydrostatic pressure in water columns increases with depth. When carried down in waters, the gas vesicles of cyanobacteria are only slightly compressed and their change in volume is negligible. Therefore, gas vesicles are effective buoyancy aids; by maintaining a certain volume, they provide a certain amount of lift to the cyanobacteria (Walsby, 1982). The next step is to show that the mechanical features of gas vesicles are also in charge of regulating the buoyancy of cyanobacteria.
Buoyancy regulation: the main function of gas vesicles
In order to float, cyanobacteria must have a lower density than that of water which is approximately 998 kg/m3 for fresh waters at 20 oC (Walsby, 1992). The density of cyanobacteria without gas vesicles ranges between 1 030 and 1 060 kg/m3 (Walsby, 1992). In this case, the formation of gas vesicles is necessary for the cell to float. A cell packed with gas vesicles in a sufficient quantity tends to have a density as low as 942 kg/m3. Here, collapsing the gas vesicles is needed for the cell to sink (Walsby, 1992). Cyanobacteria are eco strategists and possess different mechanisms to achieve this buoyancy regulation.
Gas vesicles collapse under turgor pressure
Turgor pressure is the force exerted by the gas within the vesicle on the cell walls. A normal turgor pressure equalizes the difference in water potential between the inside and the outside of the cell (Walsby, 1994). However, if increased, this force pushes the gas vesicle on the cell wall which causes stress and the collapse of the structure. Figure 12 allows a comparison of a filamentous cyanobacterium before and after the collapse of its gas vacuoles due to pressure.
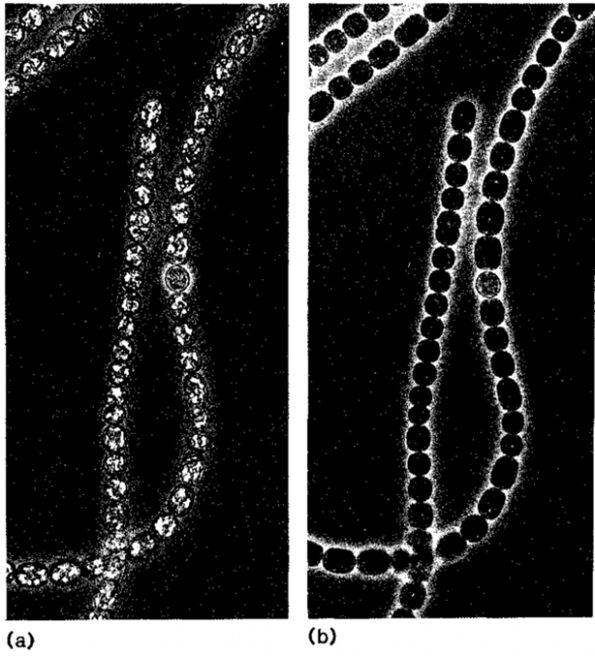
Figure 12. Filaments of Anabaena flos-aquae, a species of cyanobacteria, (a) before and (b) after collapsing due to turgor pressure. The bright refractile areas represent the gas vacuoles, aggregates of gas vesicles (Walsby, 1992).
Collapsing under turgor pressure is a buoyancy regulation method usually employed by cyanobacteria with weak gas vesicles. They tend to become overbuoyant and therefore excessively exposed to the light’s high photon irradiance. At this point, the turgor pressure surpasses the critical pressure. Then, the weaker gas vesicles collapse while the stronger ones remain which allows the cyanobacteria to sink. It takes over 16 hours of illumination for the turgor pressure to rise to 0.24 MPa (2.4 bar) and lead to a collapse of over 50 % of the gas vesicles in cyanobacteria (Walsby, 1994). This phenomenon is the first and simplest explanation of how cyanobacteria undergo buoyancy loss and become able to migrate vertically. Figure 13 demonstrates a step-by-step procedure of the collapsing of a filamentous cyanobacterium under the increase of turgor pressure.

Figure 13. The gas vacuoles of the filamentous cyanobacterium of Anabaena flos-aquae, visible under the light of a microscope and a magnitude of x 1260. The following series of photographs represent an application of increased pressure at every step from (a) to (g). Notice how the more pressure is applied, the more gas vesicles collapse, and the less distinct gas vacuoles become (Walsby, 1994).
Based on this method, there seems to be a correlation between the strength of the gas vesicles of different types of cyanobacteria and their presence in specific environments. Cyanobacteria with weaker gas vesicles are more present in shallow brine pools since they provide buoyancy more efficiently. They withstand low turgor pressures and collapse more easily. Meanwhile, cyanobacteria with stronger gas vesicles are predominant in deeper waters. The narrowness of their vesicles limits their efficiency in providing buoyancy but allows them to handle higher turgor pressures. Figure 14 uses different species of cyanobacteria to illustrate this phenomenon.
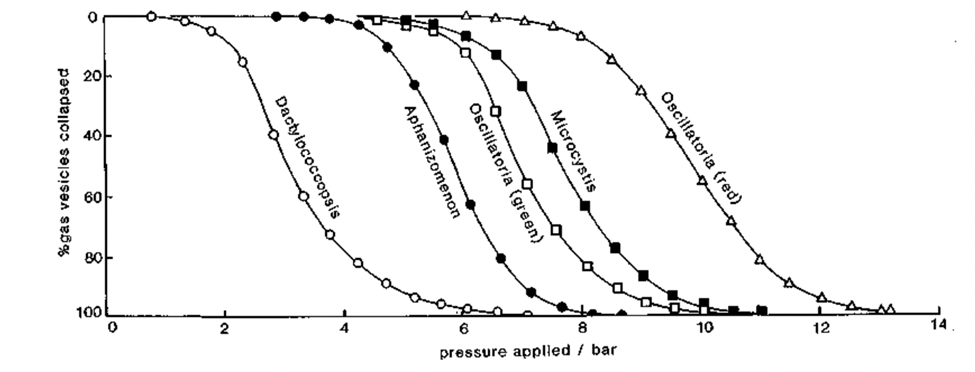
Figure 14. Pressure distributions that different types of cyanobacteria are able to withstand. Dactylococcopsis are found in shallow brine lakes and the other in freshwater lakes of ranging depths (Walsby, 1992).
Regulation of gas vesicles synthesis
After collapsing, the gas vesicles are too permeable to be reinflated. Therefore, cyanobacteria developed a kinetic regulation mechanism for the reformation of the gas vesicles Gas vesicle synthesis is not coupled to the growth and division of the cell. Starting as small bicones, it takes 6 hours for one gas vesicle to elongate and reach a length of 0.5mm (Walsby, 1992). Buoyancy recovery is a long process. The reformation of gas vesicles at a controlled rate allows the cyanobacteria to remain neutrally buoyant at stratified layers in aquatic environments. Stratification ensures the conservation of biomass while requiring minimum energy even when nutrients are limited (Reynolds, Oliver, & Walsby, 1987).
Advantages of buoyancy provided by gas vesicles
Overall, cyanobacteria use the buoyancy provided by their gas vesicles to migrate vertically up and down water columns. In natural aquatic environments, light (photon irradiance) decreases exponentially with depth due to light absorption and scattering by water (Walsby, 1994). Therefore, by floating, cyanobacteria rise to the top layers to have access to enough nutrients and the required amount of light for photosynthesis to occur and ensure their net growth. Then, they sink to retreat into deeper waters to support cyanobacterial biomass while conserving energy (Figure 15).
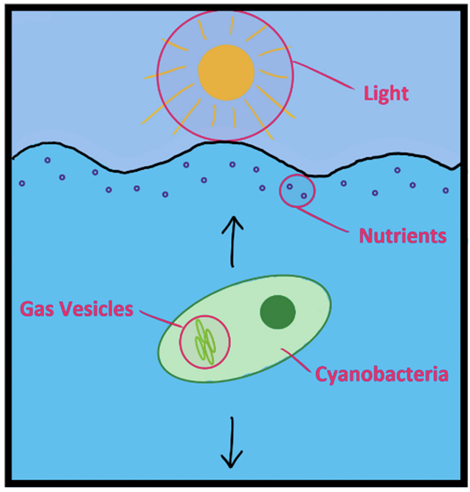
Figure 15. Simple overview of how buoyancy participates in the survival of cyanobacteria in aquatic environments. The black arrows represent the vertical up and down migrations (Hunter Hess, 2021).
Buoyancy also allows cyanobacteria to compete with other living organisms and dominate in aquatic environments. Thanks to its unique buoyancy regulation, cyanobacteria outcompete algae and other phytoplankton species by floating up to the euphotic layers and blocking their access to light. Furthermore, buoyancy is enhanced by colony formation. With a high buoyancy, the cyanobacterial colonies can persist at the surface for lengthy periods of time enabling cyanobacteria to quickly become the dominant species.
To summarize, in aquatic environments, light becomes more and more undetectable with depth and is covered by many organisms. Having access to this resource thus becomes challenging. Cyanobacteria developed different buoyancy regulation mechanisms to migrate towards top layers of water columns when light is needed and to retreat to deeper waters when enough light is absorbed. While buoyancy helps with the growth of cyanobacteria, they are not constrained to aquatic environments. The most predominant way to find cyanobacteria is in colonies, or more specifically, in biofilm.
Biofilm formation
Cyanobacteria are faced with numerous challenges presented to them by their environment, whether that be the temperature, acidity, toxicity, physical stresses such as turbulation, or other challenges. Cyanobacteria are very small and thus do not have large resistance against these stressors. So, they have evolved to use a mechanism called a biofilm to increase their survival rates.
A biofilm is a community, or colony of cyanobacteria created while attached to a surface. Mats can also be formed which are a collection of different layers of microorganism colonies that can get up to centimeters in thickness. There are a variety of factors which can influence biofilm structure including chemical, physical, and mechanical properties.
Not only does the formation of biofilm benefit cyanobacteria, but it can also benefit other microorganisms by creating optimal microenvironments in which environmental stresses are low. This can allow the proliferation of specified microorganisms that thrive only under very specific environmental conditions (Brito, 2022).
Extracellular polymeric substances (EPS)
In these communities, cells may either attach with cell-to-cell interactions or most commonly through the matrix created by extracellular polymeric substance (EPS) (Brito, 2022). The EPS is a cyanobacterial-produced substance primarily made up of polysaccharides but also proteins, lipids, and nucleic acids (Di Martino, 2018). The EPS can also take many different forms as it can be directly associated with cells, taking the form of capsular polysaccharides, or simply released into the area surrounding the cells (Brito, 2022).
The EPS plays several roles within the overall context of biofilm. Firstly, the EPS is involved in the adhesion of the biofilm to a surface (Brito, 2022). Biofilm is a sort of aqueous structure notably from the contributions of both the bacteria and the EPS (Di Martino, 2018). The cyanobacterial EPS contributes to creating a structurally stable and well-hydrated environment, as the EPS can hold onto lots of water. The EPS also contributes to cell-cell recognition, quorum sensing, (which will be explained in more detail in the chemistry essay), and horizontal gene transfer. Not only can water be retained within this complicated matrix of a substance, but nutrient storage is another feature of the EPS. Lastly, the EPS provides a physical barrier, protection to the cyanobacteria found within the biofilm. This is one of the reasons why cyanobacteria often form these communities. The physical and chemical barriers created by the EPS can protect the bacteria against stresses created by both environmental and biological sources such as light intensity, UV radiation, desiccation, salinity, erosion, antibacterial agents, and predation from protozoan organisms, or other harmful organisms in the same environment (Brito, 2022).
In bloom-forming cyanobacteria (Biofilms that are not attached to surfaces, which happens in water), the presence of adequate nutrients and light intensity results in high photosynthetic activity, leading to O2 supersaturation. This supersaturation gives rise to the formation of bubbles. The extracellular polysaccharides produced by cyanobacteria are distinctive in comparison to those of other bacteria, and they contain sulfate groups. The sulfated extracellular polysaccharides found in Synechocystis are termed “synechan.” Synechan plays a crucial role in the buoyancy and flotation of cyanobacteria cells (Mehdizadeh Allaf & Peerhossaini, 2022), as it is a molecule that allows for the oxygen to get trapped within the slimy substance that is the biofilm (Mullineaux, 2021). Within the extracellular polymeric substances (EPS) matrix, these bubbles act to elevate the biomass, causing it to accumulate at the surface. This goes back to the discussion of the buoyant forces in Equation 3. The air pockets created by the synechan have a lower density than the water surrounding the EPS, causing an upward force to occur (Mehdizadeh Allaf & Peerhossaini, 2022).
This mechanism allows the bacteria to optimize its own conditions, meaning that it can bring itself up to the surface to get more sunlight, thus producing more biomass. See Figure 16 below, which summarizes different mechanisms by which cyanobacteria can control their behaviors based on environmental conditions.
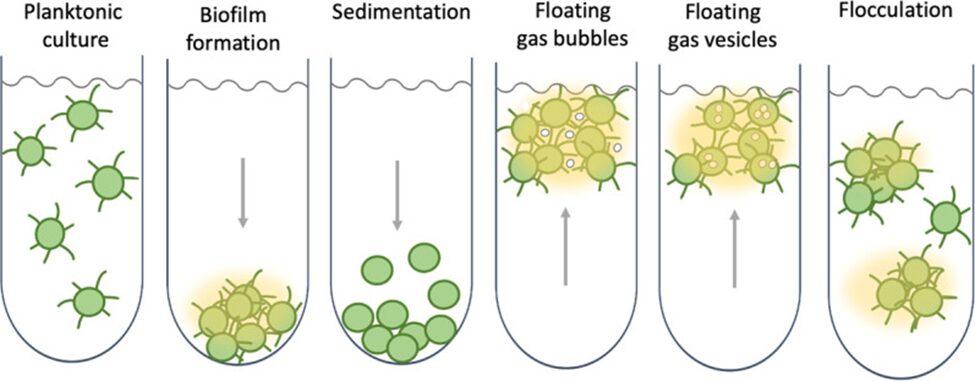
Figure 16. The various behaviours which cyanobacteria can regulate are based on environmental conditions, notably the different mechanisms of buoyancy regulation and the individual vs. grouped behaviour that the cyanobacteria may exhibit (Mullineaux, 2021).
Surface adherence and thermodynamics
Focusing on the mechanical properties of biofilm formation, cell motility plays a large role in how or whether the biofilm is created by either promoting cell-to-cell interaction or increasing dispersion forces. Looking at Synechocystis cyanobacteria as an example, a high proportion of active motile bacteria in comparison to non-motile ones results in weaker biofilm formation because of the increased dispersion forces. Non-motile cyanobacteria were therefore more trapped in the biofilm than motile varieties were. One experiment showed that soft surfaces attract more motile bacteria and that there was, therefore, a decreasing amount of biofilm over time (Mehdizadeh Allaf & Peerhossaini, 2022).
Although there are many unknown factors influencing the adherence of cyanobacteria to surfaces, one approach to determine why cyanobacteria would adhere to surfaces investigates the free energy of adherence between the bacteria and the adherent surface.
According to the Good and van Oss model (Park, 2011), The total interaction energy (∆Gadhesion), measured in mJ∙m-2 between surfaces i (microalgal/cyanobacterial cells) and I (surface materials) submerged or dissolved in water, where γlws represents the dispersive component of Lifshitz van der Waals component and √γ+s and √γ–s the polar components or the Lewis acid–base of a solid surface (Barros, 2019), is:
\Delta G_{\text{adhesion}} = \gamma_{iI}^{LW} - \gamma_{iw}^{LW} - \gamma_{Iw}^{LW} + 2 \left[ \sqrt{\gamma_w^+} \left( \sqrt{\gamma_I^-} + \sqrt{\gamma_i^-} - \sqrt{\gamma_w^-} \right) + \sqrt{\gamma_w^-} \right] \cdot \left( \sqrt{\gamma_I^+} + \sqrt{\gamma_i^+} - \sqrt{\gamma_w^+} \right) - \sqrt{\gamma_i^+ \gamma_I^-} - \sqrt{\gamma_i^- \gamma_I^+}
(5) (Barros, 2019)
The Van Oss theory divides the surface energy of solids and liquids into three parts: dispersive surface energy, surface energy due to acidic interactions (γ+), and surface energy due to basic interactions (γ−). The acidic component reflects a surface’s ability to have polar interactions with another surface that can donate electrons (act basic). On the other hand, the basic component indicates a surface’s likelihood of polar interactions with a surface that can accept electrons (act acidic). This equation relates the surface energy components, liquid components, and the contact angle, which is based on hydrophobic nature of the surface and organism (Park, 2011).
A negative ∆Gadhesion would therefore indicate favorable conditions (exothermic) to adhere, and a positive ∆Gadhesion (endothermic) would indicate unfavorable conditions for adherence (Barros, 2019).
Although a relationship in experimental data was found between the lower ∆Gadhesion and higher adherence and growth of biofilm, many discrepancies were observed, which led researchers to believe that there are other factors at play. The Van Oss (1995) approach to adhesion of microbes to surfaces overlooks hydrogen bonding between bacteria, the surface, and water, and assumes smooth surfaces with homogeneous cell distribution. In addition, although larger particles have more repulsive interactions, microalgae and cyanobacteria can reduce their effective radius, enhancing adhesion through mechanisms like EPS production and motility appendages like pili (Barros, 2019).
Despite sometimes unfavourable conditions, cyanobacteria can still adhere to surfaces by use of photosynthesis to power mechanisms in which the organism can overcome friction between the cells in the biofilm and the forces from the surfaces. For example, Synechocystis use their pili to overcome cell-cell dispersion by the motion known as twitching (Mehdizadeh Allaf & Peerhossaini, 2022). The EPS also contributes to holding the biofilm together and increasing contact between the cyanobacteria and the surface of the material (Barros, 2019).
Conclusion
In conclusion, cyanobacteria are extremely interesting micro-bacteria that have adopted several mechanisms to maintain adequate light exposure and nutrients, and to survive under harsh conditions.
Cyanobacteria live to perform photosynthesis and create biomass from carbon and light. Thus, light becomes one of the most crucial components for cyanobacterial survival. Cyanobacteria cannot control how much light the sun radiates, and so evolved optical mechanisms which allow cyanobacteria to better control light abundance. Cyanobacteria do not have the typical light sensing structure of eyes. Nature has instead developed spherical cyanobacteria with micro-scale size and shape which allow them to create a sharp image of the light source for sensitive detection based on a phenomenon similar to photonic nanojets. Filamentous cyanobacteria are also structured such that an extension of body surface area is made possible by focusing on growing length as opposed to volume. They have the opportunity to sense light gradients and have greater light exposure overall from many photoreceptors located throughout the body. This allows them to recognize light exposure and move their body to balance photosynthetic needs and possible phototoxicity depending on the light intensity and wavelength.
Finding adequate environmental conditions for survival is a related issue. Cyanobacteria cannot thrive under all conditions; thus, nature’s solution is to evolve movement mechanisms in order to move towards optimal environmental conditions. Without an obvious locomotive organelle for gliding, the force that drives cyanobacteria’s motion is generated by the secretion of adhesive slime from a junctional pore complex which allows the bacteria to move either by rotating or bending. For cyanobacteria exhibiting twitching motility, the movement is created by the extension, attachment to and object, and retraction of type IV pili. The ability to control the direction of a long-term movement incorporates various interesting mechanisms such as switching between two movement directions for P. aeruginosa. These mechanisms work in conjunction to phototaxis to maximize survival and photosynthesis.
Resource scarcity in aquatic environments is another issue cyanobacteria have to face. Nature’s solution for cyanobacteria to outcompete other organisms is the elastic compressibility of their gas vesicles. To access resources only available at the top layers of water columns, cyanobacteria control the rate of formation of their gas vesicles to float vertically. Meanwhile, to sink to the bottom and conserve energy, cyanobacteria let turgor pressure increase which leads to the collapse of their gas vesicles. In nature, the type of gas vesicles therefore depends on the depth of the aquatic environments the cyanobacteria live in. This, in turn, directly addresses the problems of resource mediation and of competition.
Lastly, nature has created a mechanism by which cyanobacteria can surmount stresses caused by extreme environments by bonding together and creating a film. Not only does the biofilm structure protect the cyanobacteria, but the molecule synechan, coupled with the EPS slime allows the capture of oxygen and thus, in an aquatic environment, causes the film to float, increasing light access.
Indeed, with billions of years of experience, cyanobacteria have become wise organisms that master all the tips and tricks to survive the conditions of constantly changing environments.
References
Biais, N., Ladoux, B., Higashi, D., So, M., & Sheetz, M. (2008). Cooperative retraction of bundled type IV pili enables nanonewton force generation. PLoS biology, 6(4), e87.
Biddanda, B. A., McMillan, A. C., Long, S. A., Snider, M. J., & Weinke, A. D. (2015). Seeking sunlight: rapid phototactic motility of filamentous mat-forming cyanobacteria optimize photosynthesis and enhance carbon burial in Lake Huron’s submerged sinkholes. Frontiers in Microbiology, 6, 930.
Brahamsha, B., & Bhaya, D. (2014). Motility in unicellular and filamentous cyanobacteria. Cell Biol. Cyanobacteria, 233-262.
Brito, Â., Mota, R., Pacheco, C. C., & Tamagnini, P. (2022). Chapter 4 – Cyanobacterial biofilms: Formation, distribution, and applications. In P. K. Singh, M. F. Fillat, V. Sitther, & A. Kumar (Eds.), Expanding Horizon of Cyanobacterial Biology (pp. 91-108). Academic Press. https://doi.org/https://doi.org/10.1016/B978-0-323-91202-0.00004-X
Brownson, J. R. S. (2014). Chapter 03 – Laws of Light. In Solar Energy Conversion Systems (pp. 41-66). Elsevier Inc. https://doi.org/10.1016/B978-0-12-397021-3.0003-X
Conradi, F. D., Mullineaux, C. W., & Wilde, A. (2020). The Role of the Cyanobacterial Type IV Pilus Machinery in Finding and Maintaining a Favourable Environment. Life (Basel), 10(11). https://doi.org/10.3390/life10110252
Daviso, E., Belenky, M., Griffin, R., & Herzfeld, J. (2013). Gas Vesicles across Kingdoms: A Comparative Solid-State Nuclear Magnetic Resonance Study. Journal of molecular microbiology and biotechnology, 23, 281-289. https://doi.org/10.1159/000351340
Di Martino, P. (2018). Extracellular polymeric substances, a key element in understanding biofilm phenotype. AIMS Microbiol, 4(2), 274-288. https://doi.org/10.3934/microbiol.2018.2.274
Ebnesajjad, S., & Khaladkar, P. R. (2005). 3 – Properties of Neat (Unfilled) and Filled Fluoropolymers. In S. Ebnesajjad & P. R. Khaladkar (Eds.), Fluoropolymers Applications in the Chemical Processing Industries (pp. 15-115). William Andrew Publishing. https://doi.org/https://doi.org/10.1016/B978-081551502-9.50006-5
Gibiansky, M. L., Conrad, J. C., Jin, F., Gordon, V. D., Motto, D. A., Mathewson, M. A., Stopka, W. G., Zelasko, D. C., Shrout, J. D., & Wong, G. C. L. (2010). Bacteria Use Type IV Pili to Walk Upright and Detach from Surfaces. Science, 330(6001), 197-197. https://doi.org/doi:10.1126/science.1194238
Havens, K. E. (2008). Cyanobacteria blooms: effects on aquatic ecosystems. Adv Exp Med Biol, 619, 733-747. https://doi.org/10.1007/978-0-387-75865-7_33
Heifetz, A., Kong, S. C., Sahakian, A. V., Taflove, A., & Backman, V. (2009). Photonic Nanojets. J Comput Theor Nanosci, 6(9), 1979-1992. https://doi.org/10.1166/jctn.2009.1254
Hoiczyk, E. (2000). Gliding motility in cyanobacteria: observations and possible explanations. Archives of microbiology, 174, 11-17.
Hoiczyk, E., & Baumeister, W. (1998). The junctional pore complex, a prokaryotic secretion organelle, is the molecular motor underlying gliding motility in cyanobacteria. Current Biology, 8(21), 1161-1168.
Hoiczyk, E., & Hansel, A. (2000). Cyanobacterial cell walls: news from an unusual prokaryotic envelope. Journal of bacteriology, 182(5), 1191-1199.
Hunter Hess, M. S., Thomas Hopkins, Ciara Hogsett. (2021). Understanding Cyanobacteria at Marion County Lake. https://www.k-state.edu/nres/capstone/NRESReportF21_TeamMoore.pdf
John F. Cochran, B. H. (2021). Archimede’s Principle. In LibreTexts Physics (Vol. 1). https://phys.libretexts.org/Bookshelves/College_Physics/College_Physics_1e_(OpenStax)
Ling, S. J., Sanny, J., & Moebs, W. (2016). Key Equations. In University Physics (Vol. 3). OpenStax. https://openstax.org/books/university-physics-volume-3/pages/2-key-equations
Maier, B., & Wong, G. C. (2015). How bacteria use type IV pili machinery on surfaces. Trends in microbiology, 23(12), 775-788.
Marathe, R., Meel, C., Schmidt, N. C., Dewenter, L., Kurre, R., Greune, L., Alexander Schmidt, M., Müller, M. J., Lipowsky, R., & Maier, B. (2014). Bacterial twitching motility is coordinated by a two-dimensional tug-of-war with directional memory. Nature communications, 5(1), 3759.
Mazard, S., Penesyan, A., Ostrowski, M., Paulsen, I. T., & Egan, S. (2016). Tiny Microbes with a Big Impact: The Role of Cyanobacteria and Their Metabolites in Shaping Our Future. Mar Drugs, 14(5). https://doi.org/10.3390/md14050097
Mehdizadeh Allaf, M., & Peerhossaini, H. (2022). Cyanobacteria: Model Microorganisms and Beyond. Microorganisms, 10(4). https://doi.org/10.3390/microorganisms10040696
Mishra, A. K., Tiwari, D., & Rai, A. N. (2018). Cyanobacteria: from basic science to applications. Academic Press.
Moore, K. A., Altus, S., Tay, J. W., Meehl, J. B., Johnson, E. B., Bortz, D. M., & Cameron, J. C. (2020). Mechanical regulation of photosynthesis in cyanobacteria. Nature Microbiology, 5(5), 757-767. https://doi.org/10.1038/s41564-020-0684-2
Mullineaux, C. W., & Wilde, A. (2021). The social life of cyanobacteria. Elife, 10. https://doi.org/10.7554/eLife.70327
Nultsch, W., Schuchart, H., & Höhl, M. (1979). Investigations on the phototactic orientation of Anabaena variabilis. Archives of microbiology, 122, 85-91.
Oliver, R. L., & Walsby, A. E. (1988). [55] Buoyancy and suspension of planktonic cyanobacteria. In Methods in Enzymology (Vol. 167, pp. 521-527). Academic Press. https://doi.org/https://doi.org/10.1016/0076-6879(88)67058-3
Park, S.-J., & Seo, M.-K. (2011). Chapter 3 – Solid-Liquid Interface. In S.-J. Park & M.-K. Seo (Eds.), Interface Science and Technology (Vol. 18, pp. 147-252). Elsevier. https://doi.org/https://doi.org/10.1016/B978-0-12-375049-5.00003-7
Pate, J. L. (1988). Gliding motility in procaryotic cells. Canadian journal of microbiology, 34(4), 459-465.
Reynolds, C. S., Oliver, R. L., & Walsby, A. E. (1987). Cyanobacterial dominance: The role of buoyancy regulation in dynamic lake environments. New Zealand Journal of Marine and Freshwater Research, 21(3), 379-390. https://doi.org/10.1080/00288330.1987.9516234
Schuergers, N., Lenn, T., Kampmann, R., Meissner, M. V., Esteves, T., Temerinac-Ott, M., Korvink, J. G., Lowe, A. R., Mullineaux, C. W., & Wilde, A. (2016). Cyanobacteria use micro-optics to sense light direction. Elife, 5, e12620.
Svercel, M., Saladin, B., van Moorsel, S. J., Wolf, S., & Bagheri, H. C. (2011). Antagonistic interactions between filamentous heterotrophs and the cyanobacterium Nostoc muscorum. BMC Res Notes, 4, 357. https://doi.org/10.1186/1756-0500-4-357
Walsby, A. E. (1982). The Elastic Compressibility of Gas Vesicles. Proceedings of the Royal Society of London. Series B, Biological Sciences, 216(1204), 355-368. http://www.jstor.org/stable/35772
Walsby, A. E. (1992). A control of gas-vacuolate cyanobacteria. In: Freshwater Biological Association.
Walsby, A. E. (1994). Gas vesicles. Microbiol Rev, 58(1), 94-144. https://doi.org/10.1128/mr.58.1.94-144.1994
Whitton, B. A., & Potts, M. (2002). Introduction to the Cyanobacteria. In B. A. Whitton & M. Potts (Eds.), The Ecology of Cyanobacteria: Their Diversity in Time and Space (pp. 1-11). Springer Netherlands. https://doi.org/10.1007/0-306-46855-7_1
Whitton, B. A., & Potts, M. (2012). Introduction to the Cyanobacteria. In B. A. Whitton (Ed.), Ecology of Cyanobacteria II: Their Diversity in Space and Time (pp. 1-13). Springer Netherlands. https://doi.org/10.1007/978-94-007-3855-3_1
Wiltbank, L. B., & Kehoe, D. M. (2019). Diverse light responses of cyanobacteria mediated by phytochrome superfamily photoreceptors. Nature Reviews Microbiology, 17(1), 37-50.