Abstract
This paper aims to obtain a better understanding of the chemical properties of Hydras. It describes the structure of the body of the Hydra as well as its behavior and interactions with its environment, with a focus on chemistry. Regeneration is a crucial part of Hydra’s survival, and the mesoglea has a vital role in this fundamental process. The chemical composition of the mesoglea is designed to support rapid tissue regeneration, highlighting the organism’s fascinating adaptability. This organism also interacts with its environment through its secretions, from its basal disk on attachment and cnidocytes on attack. The chemical compositions of these secretions will be studied in this essay. Additionally, an analysis of the animal’s innate immune system will help elucidate the physical and chemical mechanisms that protect Hydra from pathogen and help shape its beneficial microbiome.
Introduction
At first glance, this small organism may not seem likely to be one capable of surviving in an environment teeming with life. Many of the surrounding animals may consider Hydra to be a tasty snack. Recently, there have been more and more scientists interested in uncovering the secrets of Hydra, by analyzing the chemical characteristics of this animal to understand what keeps it alive. Its life cycle, defense mechanisms, and body composition all have interesting chemical properties that distinguish it from other animals, even those who belong to its same phylum! Hydra, a fascinating aquatic microorganism belonging to the class Hydrozoa and phylum Cnidaria, has long intrigued researchers due to its remarkable regenerative abilities and intricate secretory components. One of few organisms, Hydra possesses the ability to completely regenerate from small tissue fragments or even dissociated cells. Through chemical analyses, scientists have discovered the molecular mechanisms that underlie Hydra’s ability to regenerate lost body parts and create neurotoxic venom. Due to its regenerative abilities, the genus was named by Linnaeus in 1758 after the Hydra, the many-headed beast with regenerative heads in Greek mythology.
Secretory Components of Hydra
The Hydra interacts mechanically with its environment through its means of locomotion, or its detachment from a surface, as discussed in the previous essay. However, this animal also has many other interesting interactions through its secretions into the environment. We will be considering the substances that the Hydra releases to attach to a surface, as well as its secreted neurotoxins.
Basal Disk
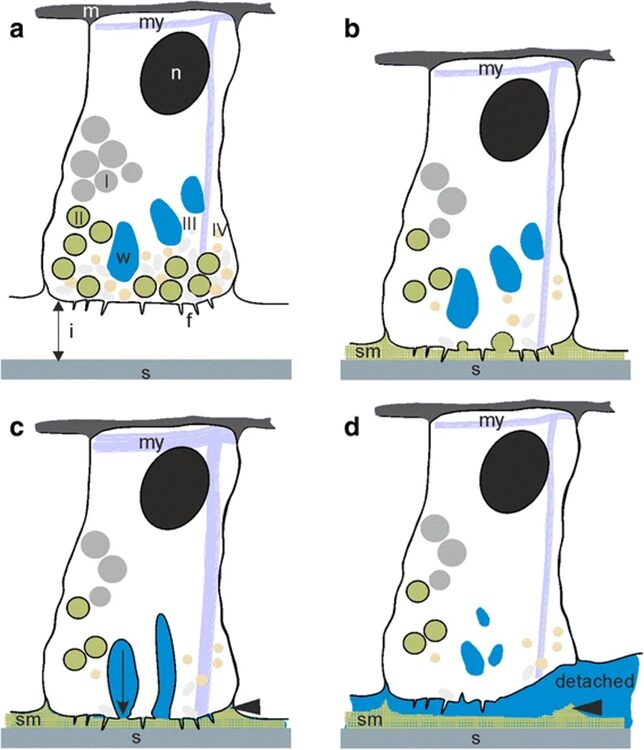
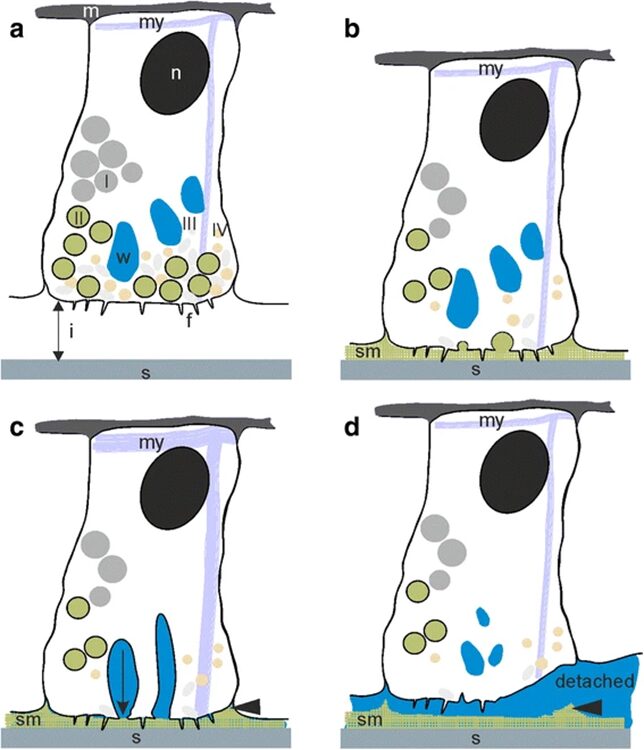
In the previous essay, we discussed Hydra’s ability to detach itself from a surface through muscular action. However, the method by which it attaches itself to surfaces is not as rooted in physics but rather, chemistry. The differences between attachment and detachment can be observed in Fig.1.
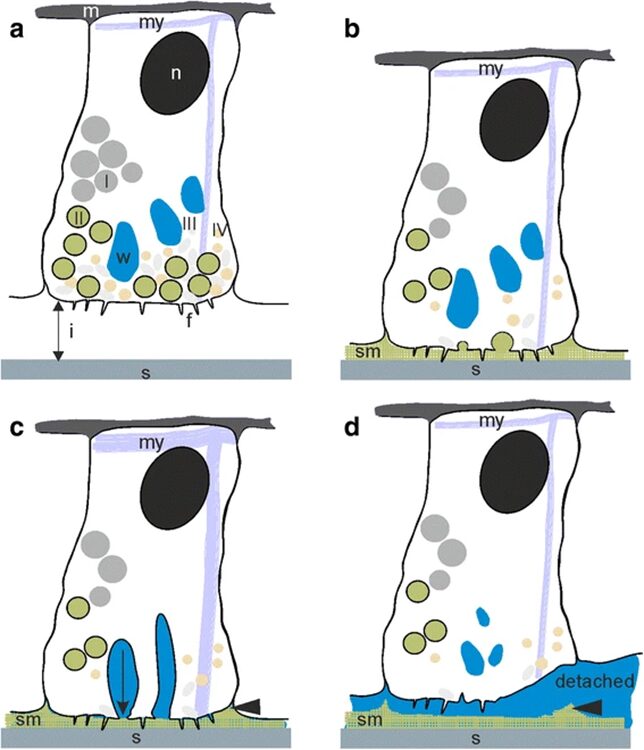
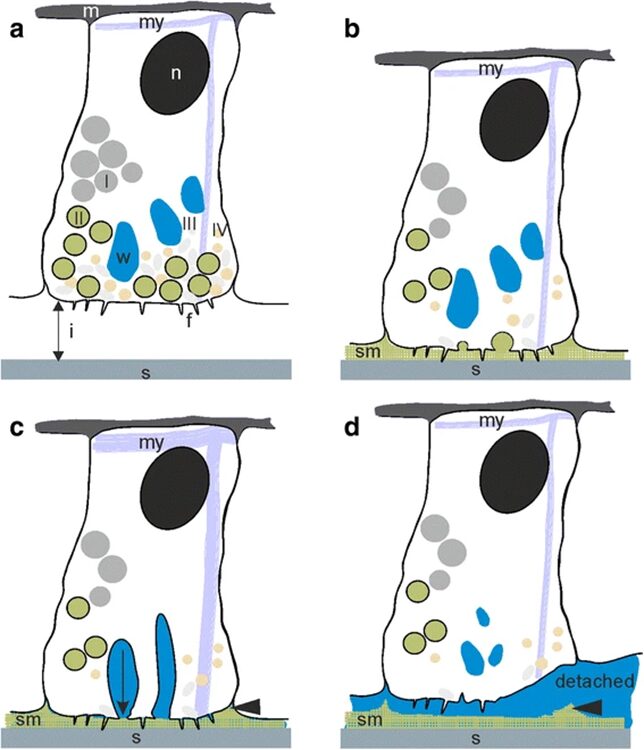
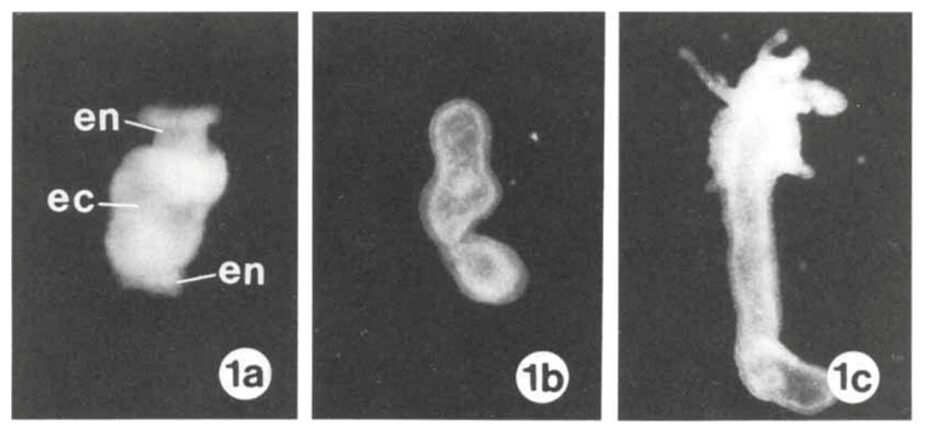
Fig. 1 Attachment by adhesion (1a, 1b) and muscular detachment (1c, 1d) (Rodrigues et al., 2016).
The figure allows us to observe the lack of a dual gland system in Hydra. It only uses a secretion to attach itself to a surface, and its muscular movement to detach. Therefore, we only need to consider the chemical composition of the adhesive material when considering the secretions by the basal disk. A 2016 paper by Rodrigues et al. studies the seven different secretory granules released by the Hydra at its basal disk during attachment to a surface and attempts to understand their roles in the adhesive properties of the secreted material (Rodrigues et al., 2016). They note that the basal disk cells share many similarities with gland cells, in that their cytoplasm contains high amounts of endoplasmic reticulum, Golgi fields and mitochondria near the storage of the secretory granules. These are important organelles in the processing and secretion of substances from the cell to its environment. Since there were no other compartmentalized adhesive components in the basal disk cells, we can conclude that these secretory granules play an important role in the adhesion of Hydra to a surface. They are secreted by the basal disk cells through exocytosis and fill the space between the Hydra’s basal disk and the substratum.
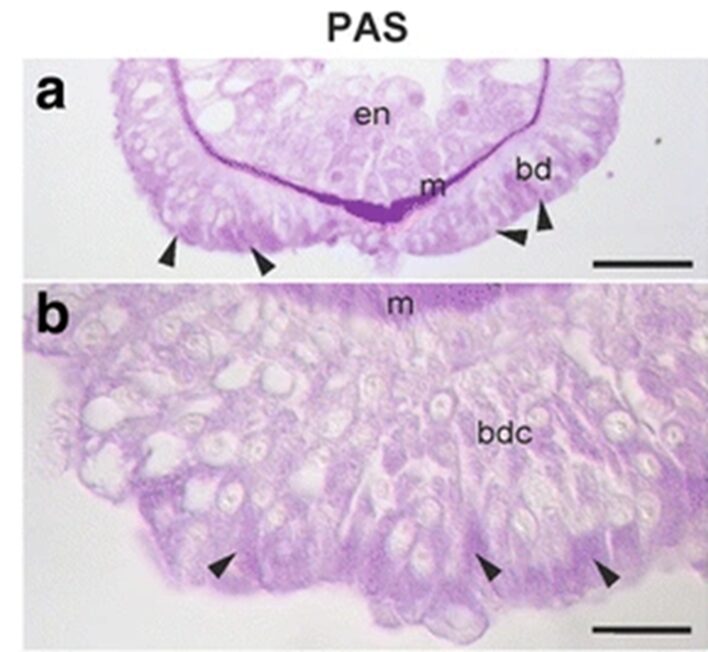
The team continued to attempt to determine the chemistry and composition of the secretory granules. It would not necessarily be useful to look to other aquatic animals that attach to surfaces, as the composition of their secretions vary greatly. For example, sea stars secrete a substance consisting of 21% proteins, 8% carbohydrates and 40% inorganic material. On the other hand, barnacles secrete a substance made of 90% protein, 1% carbohydrates, 1% lipids and 4% inorganic material. The group made use of periodic acid Schiff staining and observed the strongest reaction to take place towards the apical ends of the basal disk cells, that is where the secretory granules would be stored and secreted. We can observe this reaction in the figure below (Fig. 2).
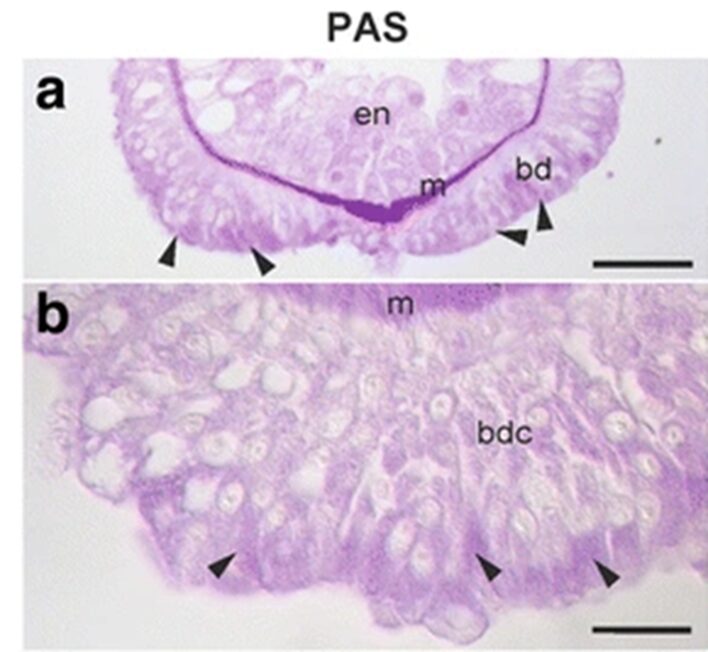
Fig. 2 PAS staining of Hydra basal disk (Rodrigues et al., 2016)
The arrows near the apical end of the basal disk indicate a strong periodic acid Schiff staining reaction. This staining method indicates the presence of mucosubstances such as mucins, glycolipids and glycoproteins. The Alcian blue staining provided a negative result for these cells, indicating that there were not negatively charged glycans in the cell for them to bind to. Therefore, they determined the presence of neutral glycans, which is odd for adhesive substances, which tend to have acidic glycans present. Both staining techniques involved submerging the Hydra in fixatives, dehydrating and embedding them into paraplast or Technovit 7100 resin. Then, they were stained with the respective liquids.
As for the lipid content of these secretory granules, this could be analysed by shining blue and green excitatory light onto the cells. The differences in the diffracted light could then be measured and used to detect the presence of lipids in these cells. Some lipid droplets were detected in the basal disk; however, they were not associated with the secretory granules. The research group with Rodrigues et al. did not provide a diagram of their data. Additionally, observation of the footprint left behind by the secretory granules revealed no lipid content, therefore suggesting that lipids are not important in the adhesion of Hydra.
The research group also used electron energy loss spectroscopy and electron spectroscopic imaging to determine the distribution of nitrogen and phosphorus in these cells. They found a relatively high density of nitrogen atoms, and some presence of phosphorus atoms. Since both play a role in the phosphorylation of proteins, this could indicate that the proteins produced for the secretory granules are post-translationally modified by phosphorylation.
Therefore, this study did not reveal the exact composition of the Hydra’s secretory granules, though it did reveal the presence of neutral glycans, the lack of lipids, and the possibility for proteins to have been post-translationally modified through phosphorylation. The Hydra’s adhesiveness is rather strong, around 66 nN. The paper draws a comparison between the sea pickle, with adhesive forces of 17 nN, and the Hydra, portraying the relative strength of the Hydra. For comparison however, a centimeter of scotch tape has a longitudinal tensile strength of 43.8N (3M, 2018). The Hydra is much less strong than this, but this makes sense, as the organism is very small and light, thus not requiring much strength to stay attached. Besides, it must be able to detach itself from the surface, which it accomplishes through its muscles.
Neurotoxins and venom
Hydrae are venomous animals, and they release neurotoxins to catch their prey. This might explain how a small animal which can hardly chase its prey has managed to survive in hostile environments surrounded by larger predators. The injection of Hydra’s venom into another organism has two main effects; a spastic paralysis and hemolysis followed by a flaccid paralysis. Spastic paralysis causes the affected organism to lose control over its movements and spasm, hemolysis results in blood cells dying and flaccid paralysis causes the organism to go limp and incapable of moving. A paper by Rachamim and Sher tells us that this second step is necessary for the Hydra to be able to ingest the prey, since it would need to be able to manipulate the organism that could be very large (Rachamim and Sher, 2012). The article also explains that there seems to be two distinct fractions in the venom, one causing the initial spasms and paralysis, and the other causing the depressing paralysis.
Some analysis of the toxins present in Hydra’s venom has revealed resemblances to at least eight commonly known toxins. Actinoporins, for example, are found in sea anemones and are lethal proteins responsible for attacking surrounding prey. The group also found proteins containing specific sequences of amino acids called ShK domains. It is theorized that these may block potassium ion channels (Rangaraju et al., 2010). This could have adverse effects on a cell, as these channels are essential for the cell to maintain its gradient of electrical potential. A depolarized cell would fail to communicate through electrical signals, which may cause paralysis of the organism.
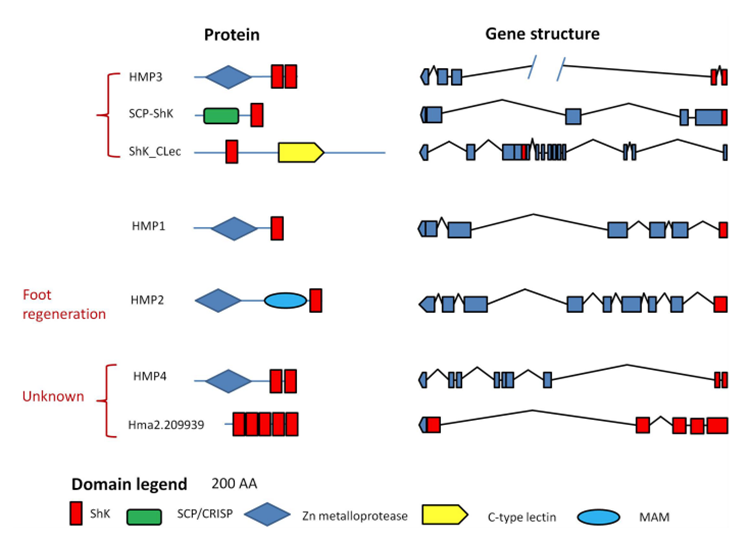
Hydrae are similar to sea anemones, cone snails and scorpions since they all must paralyze their prey immediately in order to kill and consume it; however, Hydrae’s venom is different from those of these other animals. Hydrae’s venom has not been observed to contain any short peptide neurotoxins, which is what these other animals mainly use in order to paralyze their prey. Instead, the Hydra mainly rely on the ShK domain containing proteins and some neurotoxic PLA2s. The presence of this ShK domain can be seen in Fig.2, which demonstrates the different domains present in the proteins that make up Hydra’s neurotoxins.
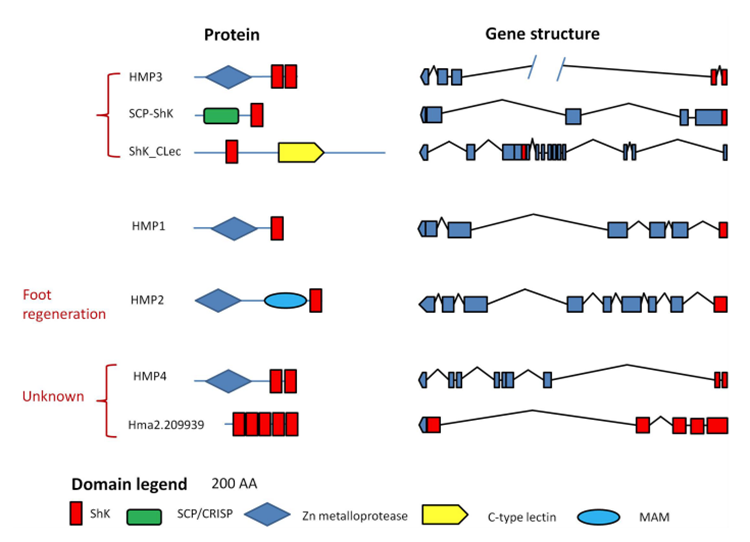
Fig. 3 Domains found in proteins from Hydra neurotoxins (Rachamin and Sher, 2012).
There are also many other toxic and non-toxic components that work together to induce a desired effect in the Hydra’s prey. The article discusses that the effect probably does not arise from a sum of all the effects caused by the different components, but that they somehow synergize to achieve the effects. At the molecular level, they may allosterically modify their binding to a molecular target. They also interact at the cellular level by affecting multiple complementary targets in nerve or muscle fibers. The example they provide is opening sodium ion channels and blocking potassium ion channels, which would lead to very rapid depolarization and send overwhelming signals through the target organism. The different components of the venom can also act at the level of systems; where they could affect many critical systems at once, leaving the prey with many problems that it would need to address before resuming normal function. This makes it very difficult for the prey, the live water flea (Loomis, 1959), for example, to defend itself against the Hydra.
The researchers also question how Hydrae may affect their surrounding environment through their neurotoxins. This is because the neurotoxins are not necessarily only delivered directly into the Hydra’s prey through its nematocysts, but also contains hormones that regulate other instances of releasing these neurotoxins. More specifically, a released neurotoxin into an animal from a cnidocyte should reduce the likelihood of other cnidocytes also releasing their neurotoxins. This would likely be done to prevent waste from excessive delivery of neurotoxin. This hormone should also signal to the Hydra that it should prepare to feed on the prey that it has just neutralized. This internal communication through hormones suggests that communication may occur on other scales, for example between different Hydrae. This may be likely considering that Hydrae tend to live in clusters, and communicating could promote group response to many preys arriving at the same time, for example. If Hydrae signal between themselves using their venom or some other chemical components, it is possible that their prey can detect these signals. Therefore, Hydrae may be altering the chemical landscape on the micron scale on which Hydrae interact with other bacteria and animals.
Mesoglea of Hydra
Chemical Composition
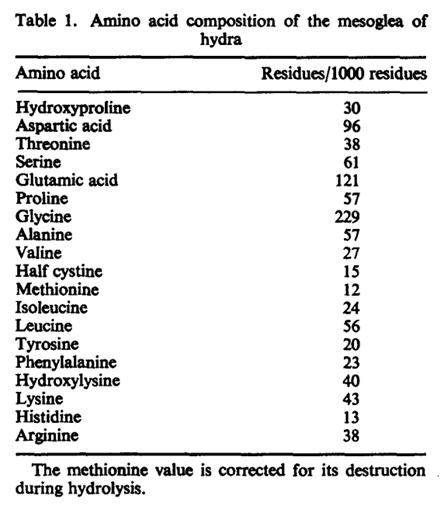
The mesoglea serves as a basement membrane for the two epithelial layers of Hydra. In addition, it is hypothesized to play other morphogenetic roles, perhaps directing cell migration or regulating the regenerated form in hydra (Epp et al., 1986). The membrane is chemically composed of neutral sugars and amino acids. This composition was determined following an isolation method of the mesoglea which required repeated freezing and thawing in detergent and treatment with disulfide reducing agent to remove adhering nematocysts (Barzansky & Lenhoff, 1974). The process had no contaminants, and a high amount of neutral sugars was found in Hydra including glucose and galactose. Small amounts of fucose, rhamnose, and amino sugar glucosamine were also detected. From the chemical breakdown, researchers were able to determine that the major protein component of the mesoglea is collagen (Barzansky & Lenhoff, 1974). This was found through detection of hydroxylysine, hydroxyproline, proline, and large amounts of glycine which make up the chemical structure of collagen as shown in Fig. 3. Glucose and galactose also join as a dimer by an alkali stable bond to make up the polypeptide backbone of collagen. From the composition, it was determined that chemical composition of mesoglea from hydra resembles collagen of vertebrate basement membranes more than “extractable” vertebrate collagens. The main distinction between the two collagens is that “extractable” vertebrate collagens contain less than 10 residues per thousand of hydroxylysine while vertebrate basement membranes contain over 20 residues per thousand of hydroxylysine. The Hydra mesoglea contains 30 residues per thousand hydroxylysine as shown in Fig. 4. The vertebrate basement membrane is mainly composed of type IV collagen and two glycoproteins. Hydra’s mesoglea was also found to be composed of 6.7% in neutral sugars and 1.4% in glucosamine in dry weight (Barzansky & Lenhoff, 1974). The large sugars were detected using thin layer chromatography and suggest that the mesoglea contains a disaccharide unit like collagens. Both the mesoglea and vertebrate basement membranes contain a significantly greater amount of covalently linked neutral sugars than extractable vertebrate collagens. Extractable vertebrate collagens contain less than 2% of dry weight in neutral sugars while vertebrate basement membranes contain glycoproteins and neutral sugars.
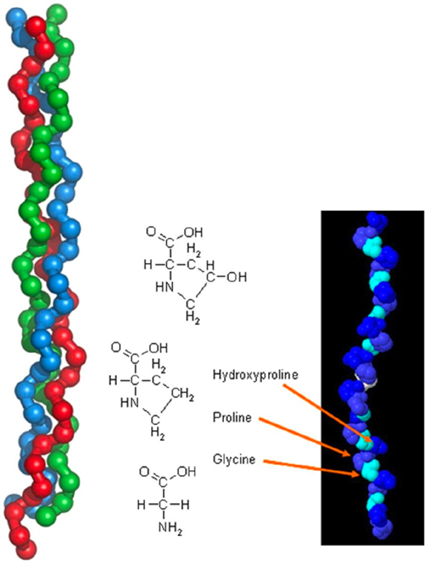
Fig. 3. General chemical structure of collagen (left) and tropocollagen (right) (Rau & Kajzar, 2015).
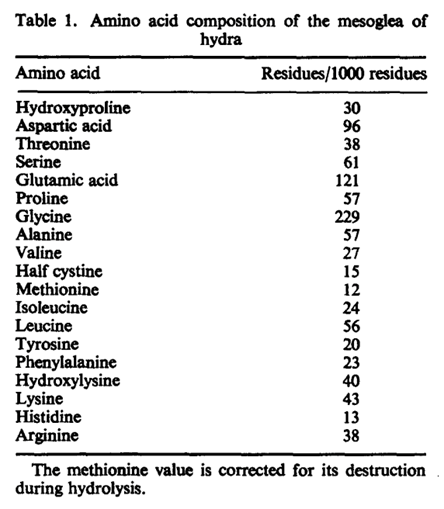
Fig. 4. Amino acid composition of mesoglea of hydra (Barzansky, 1972).
In order to confirm that the Hydra mesoglea is most similar to the vertebrate basement membrane, researchers performed in-situ immunocytochemical analysis procedures at light and electron microscopic levels. With the use of an antiserum, researchers were able to test for various antibodies to ECM components such as type IV collagen, laminin, fibronectin, and heparan sulfate proteoglycan in vertebrate ECM membranes. As shown in Fig. 5., the mesoglea reacted positively with all antibodies tested, indicating the strong similarities mesoglea have to vertebrate membranes (Sarras Jr et al., 1991). As a result, Hydra mesoglea is formed from glycoproteins of collagen resembling the vertebrate basement membrane in composition. The studies also found that the matrix molecules were evenly distributed across the mesoglea, as seen in vertebrate basement membranes. This result indicates that there is also a degree of similarity in the structural and functional properties of the two ECMs. The immunocytochemical studies also illustrated a fluorescent pattern in the ectodermal layer, indicating that the epithelial layer has a prominent biosynthetic role in mesoglea formation.

Fig. 5. Reactivity of Mesoglea with antibodies to vertebrate membrane ECM components (Sarras Jr et al., 1991).
Role in Morphogenesis
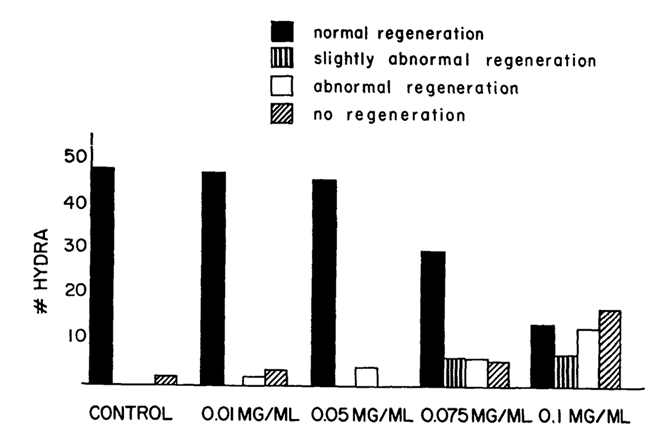
An essential function of vertebrate basement membranes is their role in morphogenesis. Since the mesoglea of Hydra resembles the vertebrate basement membrane in composition, there is a possibility it has a role in morphogenesis as a contributing factor to the establishment of a regenerated form of adhering cells. This theory was tested using lathyrogen 3-aminopropionitrile (BAPN), an inhibitor of collagen crosslinking. The inhibitor causes abnormalities in vertebrate connective tissues by binding to and inhibiting enzyme lysyl oxidase which catalyzes the first step of cross-linking of collagen. By using this inhibitor, researchers were able to discover if new cross-linked mesoglea is required for normal regeneration to occur (Barzansky & Lenhoff, 1974). The method began with removing the head, just below the hypostome, of the Hydrae and placing the bodies in varying concentrations of BAPN. The Hydra bodies were left to sit in the solution and observed for over 72 hours. As shown in Fig. 6., the higher the concentration of BAPN, the higher the number of abnormal regenerations. Additionally, the longer the Hydra was left in the BAPN solution, the more abnormal the regeneration. The blockage of head regeneration was reversible, indicating that it was not due to a nonspecific toxic effect of the drugs. Through this experiment, it was also found that Hydra placed in BAPN between 24-48 hours after head removal, regenerated like Hydra in BAPN for 72 hours. This indicates that an important cross-linking event occurs between 24-48 hours after removal for the regeneration process. Morphogenesis is a critical component of Hydra’s survival. Without proper synthesis of the mesoglea, Hydra are unable to perform regeneration which puts their survival at risk. From this, researchers were able to conclude that alterations to the mesoglea affect the overall regeneration of the cell shape.
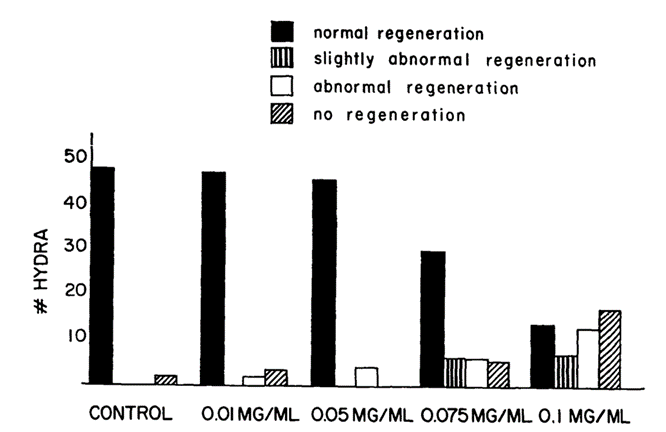
Fig. 6. Results of varying concentrations of BAPN used to test hydra regeneration, scored 48 hours after head removal. 50 Hydrae used for each condition (Barzansky & Lenhoff, 1974).
To investigate the synthesis and structure of the mesoglea, the separation and recombination of the ectoderm and endoderm layers was performed. The head and foot of the Hydra were removed, and the body was soaked in a solution to properly separate the layers. The endoderm was then reinserted in the ectoderm and left to culture and regenerate for several days. This process provides insight on the morphology of the mesoglea through the use of light and electron micrographs. Early mechanisms of hydra cell aggregate formation are based on cell sorting events involving adhesive properties of the epithelial cells (Technau & Holstein, 1992). Once the ectoderm is established, no further rearrangement occurs. During regeneration, it was found that the mesoglea disappears 6-8 hours after reassembly and a hollow regenerate forms. The “new” mesoglea undergoes synthesis by hour 12 after reassembly (Epp et al., 1986). The mesoglea has a trilaminar appearance after undergoing synthesis which suggests that it originates from both epithelial layers, the ectoderm and endoderm. The mesoglea functions in Hydra as a basement membrane, providing structural support for the two epithelia since it is a site of contact between the two cellular layers. Thus, it is responsible for intercellular communication between the epithelia. After 48 hours past reassembly, the mesoglea returns to its normal appearance. Once 7 days have passed, the Hydra reaches its final regenerated form as shown in Fig. 7. Mesoglea precursors were injected as an amino acid mixture into the mesoglea. However, 5-6 hours were required for the newly acquired amino acids to be incorporated in the mesoglea. The turnover of amino acids is slow and it should be noted that after tissue reassembly, free amino acids do not travel from the endoderm to ectoderm and are therefore not utilized by the ectoderm (Epp et al., 1986). Despite Hydra’s classification as an early divergent metazoan group and a lower animal form, their development of a fundamental tissue pattern adherent to the ECM is utilized in higher organisms. In terms of evolution, the Hydra mesoglea is one of the earliest forms of ECM to have defined tissue layers (Sarras Jr et al., 1991).
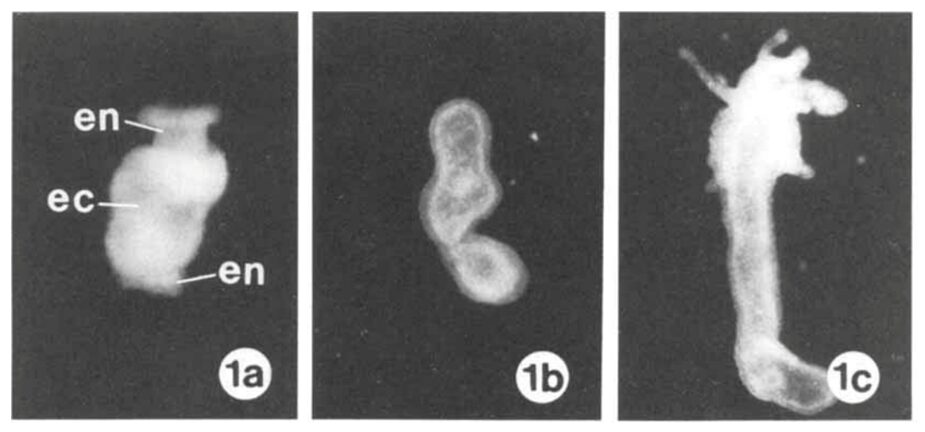
Fig. 7. (1a) Mesoglea immediately after reassembly of endoderm (en) and ectoderm (ec). (1b) After 6-8 hours, hollow regenerate forms. (1c) After 7 days, a regenerated Hydra is formed (Epp et al., 1986).
A Hydra organism is able to reorganize and regenerate its adult structure within 96 hours to 7 days from dissociated Hydra cells as shown in Fig. 8. Researchers discovered that hydra cell aggregates form an epithelial later by hour 12 of development and follow by developing the mesoglea. Once the mature mesoglea has formed by 48-72 hours of development, morphogenesis of Hydra begins (Sarras Jr et al., 1993). This indicates that if mesoglea formation does not occur, the morphogenesis of a Hydra cell is blocked. The reason for this is that as Hydra cell shapes change during morphogenesis, the contact between the cell and substrate varies. If the mesoglea is not available in sufficient amounts to accommodate the cell contact, the shape change will affect the morphogenesis (Barzansky & Lenhoff, 1974). Additionally, the morphogenesis process involves the establishment of activation and inhibition gradients for head formation. As the head-activation potential increases, the rate of morphogenesis of the head increases (Sato & Sawada, 1989). The remarkable ability of Hydra to reorganize and regenerate its adult structure within a short timeframe showcases the intricate design of nature. This regeneration process, marked by the critical formation of the mesoglea, underscores the essential role the mesoglea plays in Hydra’s morphogenesis.
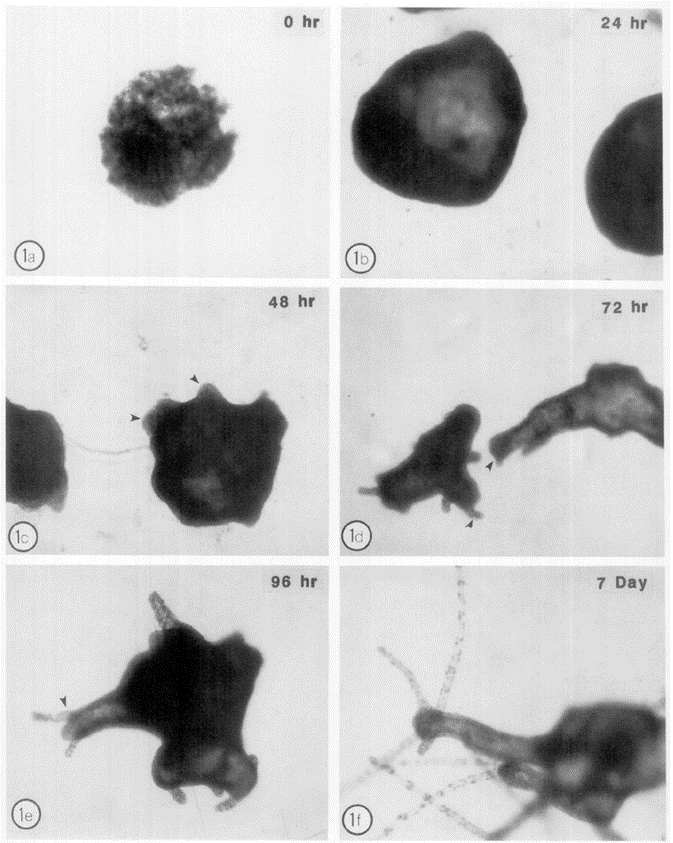
Fig. 8. Morphogenesis pattern of dissociated hydra cell aggregates through 7 days of development (Sarras Jr et al., 1993).
The innate immune system of Hydra
In vertebrates, the immune system is separated in two parts: a fast-responding innate immune system that is limited in its effectiveness and a slow but efficient adaptive immune system that evolves as the organism grows. However, invertebrates such as Hydra lack that adaptive immune system as well as mobile phagocytes, hemolymph and impermeable barriers from cuticles or an exoskeleton (Bosch et al., 2009). Instead, they rely entirely on their innate immune system to defend itself against bacterial infections in everyday life.
The role of Hydra epithelia in immunity
Hydra is a soft-bodied animal with two monolayered epithelia, the outside ectoderm, and the inner endoderm inside the gastral cavity. To guard against the outside environment, the ectoderm is covered by multiple layers of glycocalyx (Schröder & Bosch, 2016).
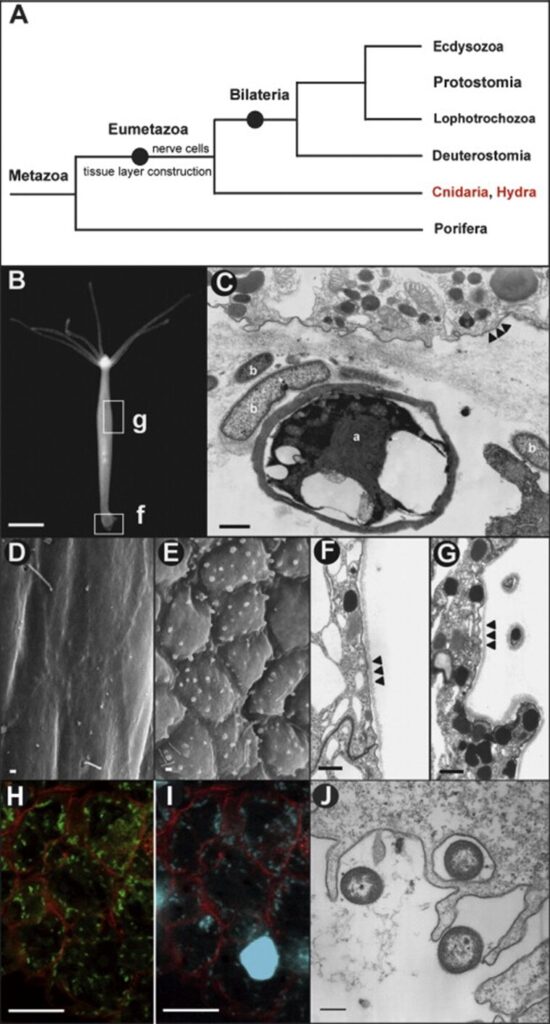
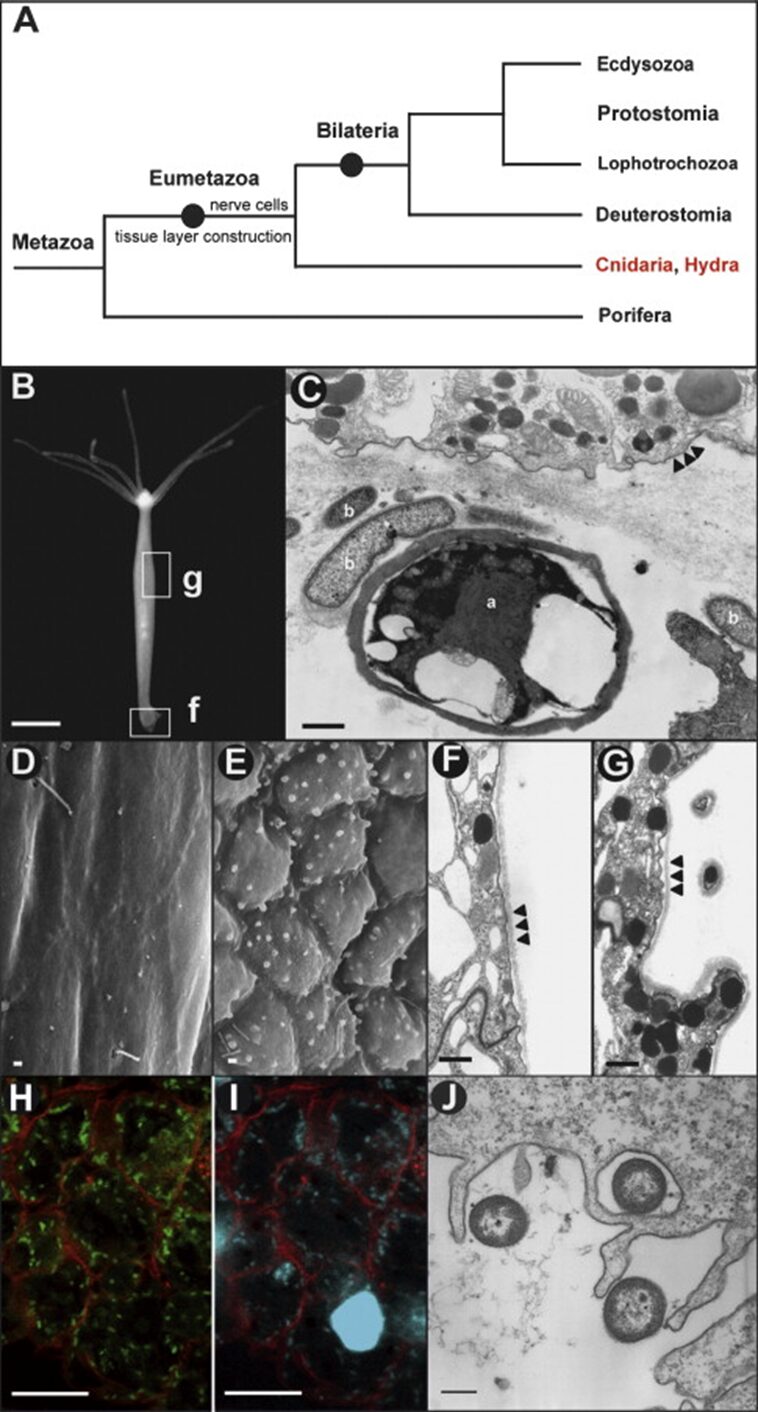
Fig. 9. Hydra and the epithelial cellular response to pathogens. (A) Phylogenetic tree of Cnidarians. (B) Hydra magnipapillata; g, gastric region examined by reflection electron microscopy in (D-G); f, foot examined by transmission electron microscope in (C). (C) foot epithelial cells exposed to microbes; a, alga; b, bacteria. Arrowheads show cell membranes. (D,F) REM and TEM showing ectoderm in control Hydra magnipapillata polyps. Arrowheads show outer cell membrane and glycocalyx. (E,G) REM and TEM showing ectoderm in polyps exposed to filtrates of Pseudomonas aeruginosa. Arrowheads show outer cell membrane and glycocalyx. (H-J) Microscope analysis of bacteria in endoderm. (H) CLSM showing bacteria expressing GFP in endoderm engulfed during food intake. (I) Cells in (H) stained with Hoechst. (J) TEM showing endodermal epithelial cells engulfing bacteria (Bosch et al., 2009).
Embedded in the glycocalyx are diverse numbers of transmembrane glycoproteins, proteoglycans and glycolipids that are constantly renewed. As such, the glycocalyx plays a crucial role in cell-cell recognition and receptor-ligand interactions as well as protecting the monolayered ectoderm (Schröder & Bosch, 2016). More precisely, in Hydra, the glycocalyx is 1.5 µm thick and can be separated into five morphologically distinct layers, with the outermost layer accounting for more than half of the structure. Despite its size, the outer layer is a loose meshwork that can be easily washed off with a high concentration salt solution. Conversely, the inner four layers are densely packed and attached to the surface of the ectoderm (Schröder & Bosch, 2016). As such, the mucus-like outer layer of the glycocalyx acts not only as a renewable buffer region, but also as a habitat for a symbiotic bacterial community, while the inner four layers served as a physicochemical barrier, given that no bacteria were observed entering the inner layers (Schröder & Bosch, 2016). Thus, the glycocalyx is an ingenious border that maintains itself and gives refuge to beneficial bacteria. In addition, when exposed to bacterial pathogen-associated molecular patterns (PAMPs) from Pseudomonas aeruginosa, the ectodermal epithelial cells changed morphology (Fig. 9D-G) and formed round blebs at the surface, which, through an analysis of the thin sections (Fig. 9G), showed a high content of granules. Consequently, Hydra epithelial cells exhibit cytoskeleton rearrangement and increased secretion when exposed to PAMPs (Bosch et al., 2009; Schröder & Bosch, 2016). Comparatively, the endodermal epithelium lacks any glycocalyx despite being exposed to numerous bacteria and pathogens from ingesting prey (Fig. 9H-J). As such the endodermal cells compensate for the lack of physical barrier by producing antimicrobial peptides (Bosch et al., 2009).
Antimicrobial peptides (AMP) in Hydra endoderm
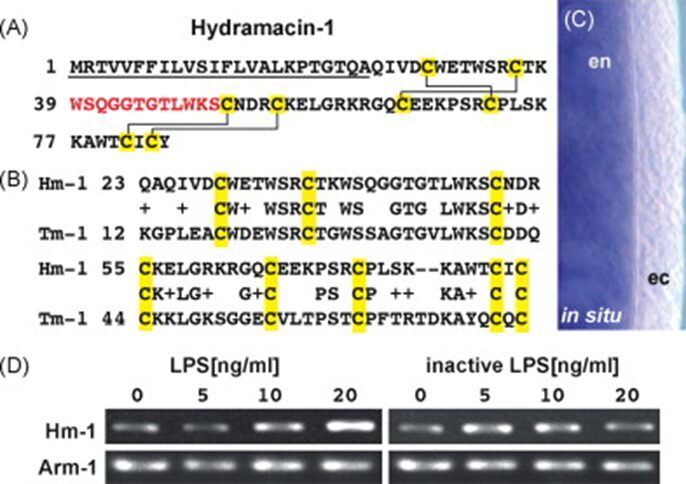
By exposing Hydra polyps to P. aeruginosa culture filtrate, an innate immune inductor composed of lipopolysaccharides and flagellin, researchers were able to stimulate and identify an unknown Hydra AMP called Hydramacin-1(Bosch et al., 2009). The cDNA of Hydramacin-1 encoded a cationic 60 aa peptide that contained eight cysteines and had a monoisotopic molecular mass of 7009 Da (for a peptide with four cysteine-bridges) (Fig. 10A). The fact that the natural Hydramacin-1 has a lower mass of 15 Da shows that a molecule of NH3 was removed and two hydrogens were added during post-translational modification, signifying that natural Hydramacin-1 had a pyroglutamate at its N-terminus (Bosch et al., 2009). Equally, in situ hybridization demonstrated that the Hydramacin-1 mRNA is expressed solely in the endoderm (Fig. 10C) (Bosch et al., 2009). Since the ectodermal epithelium has a glycocalyx, it is an efficient use of resource to limit Hydramacin expression to where it is most needed. Similarly, when determining whether this AMP is continuously produced or is induced by pathogen products, Hydrae were exposed to isolated liposaccharides at varying concentrations (Fig. 10D) and the results showed that Hydramacin-1 expression was induced by PAMPs and was dose-dependent rather than constitutively expressed (Bosch et al., 2009).
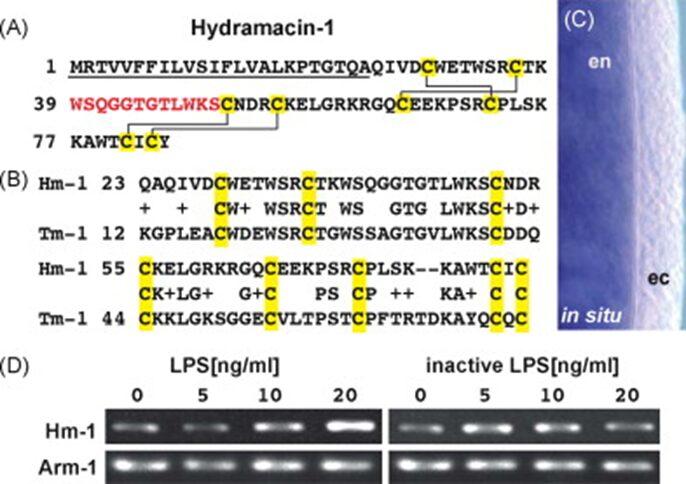
Fig. 10. Hydramacin-1 gene in Hydrae. (A) Hydramacin-1 amino acid sequence. Underlined is a predicted signal peptide followed by a cationic region. Highlighted in yellow are cysteines that form disulfide bridges indicated in solid lines. In red are the amino acids found by mass spectrometry. (B) Comparison with theromacin from Aplysia californica. (C) In situ hybridization to show exclusive expression in endoderm. (D) Ultrapure lipopolysaccharide with hydra to show dose-dependent induction of AMP gene expression (Bosch et al., 2009).
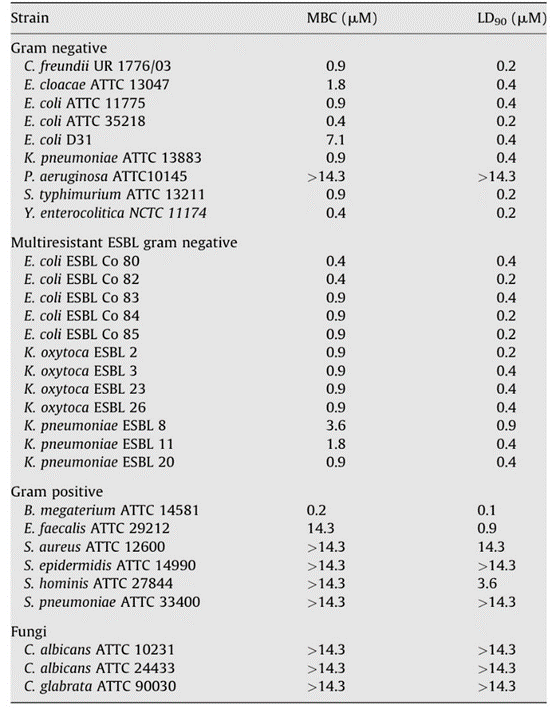
Researchers were then able to test the antimicrobial capabilities of Hydramacin-1 by using an E. coli fusion protein and forming a recombinant rHydramacin-1 peptide. The resulting protein inhibited the growth of B. megaterium in liquid growth inhibition assays as well as other bacteria, some of which are antibiotic-resistant human gram-negative pathogens (Fig. 11) (Bosch et al., 2009). However, Hydramacin-1 proved to be ineffective against gram positive Coccus species and gram-negative non-fermentation species, as well as P. aeruginosa. Given that P. aeruginosa is a pathogen associated with Hydra and that its filtrate products were used to induce antimicrobial response in Hydra, leading to the expression of Hydramacin-1, it is surprising that Hydramacin is unable to inhibit this bacterium. This is subsequently explained by the fact that the supernatant also contained LPS, which does strongly induce Hydramacin-1. In contrast, against a bacterium with few LPS structures in its outer membrane such as E. coli strain D31 (Fig. 11), Hydramacin is also shown to have reduced activity (Bosch et al., 2009). Lastly, while Hydramacin is unable to deal with P. aeruginosa, Hydra endodermal epithelial cells also produce other AMPs from the periculin and arminin peptide families that are likely able to counter it (Bosch et al., 2009).
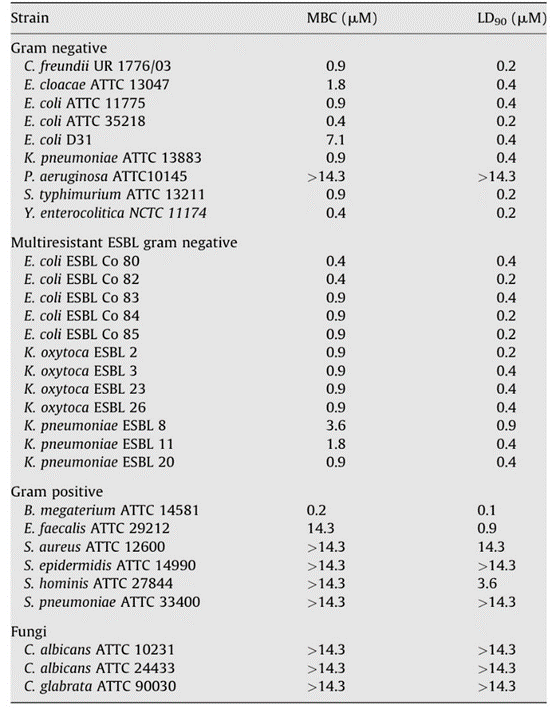
Fig. 11. Table showing antimicrobial activity of Hydramacin (Bosch et al., 2009).
Consequently, antimicrobial peptides play an essential role in ensuring the security of the endodermal epithelium despite its lack of a physical barrier, considering that such a barrier in the gastric cavity would hinder digestion. However, AMP production, being dose-dependent, suggests the presence of a receptor that recognises PAMPs and induces an antimicrobial response.
Transmembrane receptors responsible for bacterial recognition
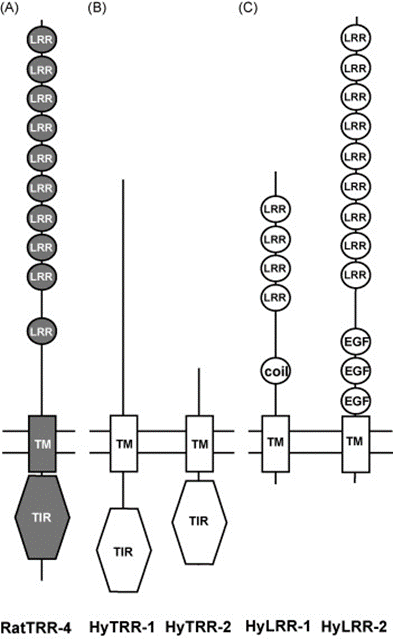
Toll-like receptors (TLR) are protein receptors that identify common microbial components and are vital to the innate immune response as they serve to detect pathogens and distinguish them from symbiotic bacteria. They can be found in various animals and have been conserved throughout evolution (Bosch, 2014). In Hydra, researchers searched the Database of Expressed Sequence Tags and identified two genes that form structures similar to TLR proteins in vertebrates with a Toll/interleukin-1 receptor domain (TIR), a transmembrane domain and an extracellular domain (Fig. 12B). These two genes have been named Toll-receptor-related 1 and 2 (HyTRR-1 and HyTRR-2) (Bosch et al., 2009). However, unlike TLR, HyTRR-1 and HyTRR-2 lack leucine-rich repeats (LRR) in the extracellular domain. Instead, using human LRR sequences as a reference, researchers found two other genes that encode transmembrane proteins carrying TLR-related LRRs in the extracellular domain, called HyLRR-1 and HyLRR-2 (Fig. 12C) (Bosch et al., 2009).
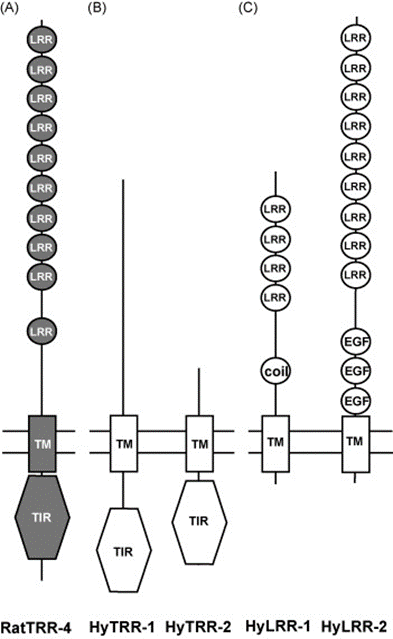
Fig. 12. TLR-like receptors in the innate immune system in Hydra. (A) Rat TLR-4 showing TIR domain in cytoplasm, TM domain and extracellular domain with LRRs. (B) HyTRR-1 and HyTRR-2. (C) HyLRR-1 and HyLRR-2 (Bosch et al., 2009).
To determine whether the extracellular HyLRR-2 protein, which resembles vertebrate TLRs the most, can recognize PAMPs, the TIR domain of IL1R1 was fused with HyLRR-2. This formed an artificial transmembrane vertebrate-like TLR protein, which was able to detect bacterial flagellin in human HEK293 cells(Bosch et al., 2009). Furthermore, dsRNA-mediated interference (RNAi) preventing HyLRR-2 transcription in Hydra caused not only a depletion of HyLRR-2 transcripts, but also of HyTRR-1 transcripts and vice versa, suggesting a relationship between the two genes. Equally, both HyTRR-1 and HyLRR-2 knockdowns reduced the expression of Periculin-1, demonstrating that the Periculin-1 gene expression is downstream of HyLRR-2 signal cascade (Bosch et al., 2009). This follows the logic that it is the recognition of PAMPs that causes the AMPs to be produced as an immune response. Further investigation of the relationships between TLR-like receptors in Hydra revealed that HyLRR-2, acting as receptor, and HyTRR-1, as signal transducer, interacted and worked together (Fig. 13) to recognize PAMP flagellin and trigger an innate immune response, causing the production of AMPs (Bosch, 2014; Bosch et al., 2009).
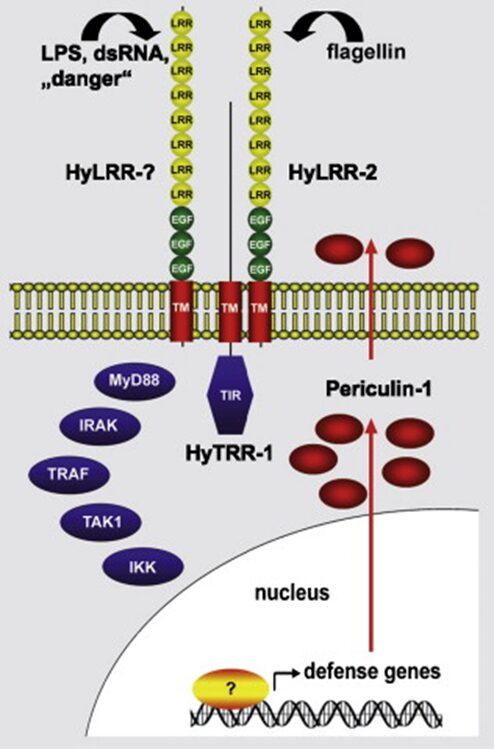
Fig. 13. Model of components and pathways involved in the Hydra innate immune system. Microbial components (LPS, flagellin), viral components (dsRNA) and danger signals are recognized by HyLRR-2 transmembrane proteins in the epithelial cell membranes. A signal is then triggered and transduced by HyTRR-1 or-2, causing a cascading reaction. This activates AMP gene expression and produces AMP like periculin-1, which are secreted outside to eradicate foreign bacteria (Bosch et al., 2009).
Ultimately, from the mucus-like glycocalyx that can house symbiotic bacteria, to the TLR-like receptors that induces complex AMPs capable of inhibiting even antibacterial-resistant human pathogens, Hydrae evolved a complex and capable innate immune system despite its morphological simplicity. Incredibly, each component of this system serves a specific purpose that compensates for the weaknesses found elsewhere. Indeed, the glycocalyx provides badly needed cover that always protects and encompasses the entire exterior epithelium. Comparatively, the inner endodermal epithelium is already sheltered and therefore serves as the medium for digestion. However, without glycocalyx, it is also more vulnerable to the occasional pathogen that does slip through. Consequently, powerful bactericides capable of eliminating dangerous pathogens are produced (AMP). Additionally, producing AMP continuously is resource draining and unnecessary. HyTRR-1 and HyLRR-2 receptors consequently allow for the induction of these AMPs when needed, allowing beneficial bacteria to proliferate at ease while keeping harmful elements out. As such, all these mechanisms have adapted to perfectly suit the needs of the organism, while protecting it as best as possible and in an efficient manner, which is why they have been conserved throughout evolution. Indeed, the mucus layers and AMP effectors can be found in vertebrate gut and TLR is present in human macrophages (Schröder & Bosch, 2016). Lastly, another key reason behind the Hydra innate immune system’s effectiveness is its ability to manage a diverse symbiotic microbial community that benefits both the bacteria and the host (Bosch, 2014).
Hydra as a Metaorganism
As seen in the previous section, Hydra’s innate immune system, with its host-specific antimicrobial peptides (AMPs) and microbial recognition signaling pathways, has evolved to fight against pathogens. However, by considering Hydrae as a metaorganism (a metaorganism refers to a macroscopic host and its synergistic interdependence with other prokaryotes or eukaryotes), its innate immune system also plays a crucial role in detecting and controlling resident beneficial microbes. In a study, two Hydra species displayed significantly different microbiota that were similar to that of the respective species in the wild, despite being kept in the same lab for more than 20 years under identical conditions. This discovery provides strong evidence that each host species actively selects their own bacterial community through genetic encoding in the epithelium (Fraune & Bosch, 2010). But what is the benefit of this microbiota selective pressure? Through what mechanisms are they selected? These are the questions this section will aim to answer, and we will begin by taking a closer look at the mode of action of antimicrobial peptides.
Antimicrobial peptides
Antimicrobial peptides (AMPs) are short and generally positively charged peptides found in a wide variety of life forms. Their number of amino acid residues is between 10 and 60 (average: 33.26), and almost all AMPS are cationic (average net charge: 3.32) (Huan et al., 2020). Since AMPs have complex mechanisms of action and do not readily lead to the development of resistance, designing synthetic AMPs based on natural templates garners significant research interest currently in the context of emerging antibiotic-resistance and declining discovery of new antibiotics (Huan et al., 2020). AMPs’ mechanisms of action can be divided into membrane targeting mechanisms, described through various models detailed below, or non-membrane targeting mechanisms including the inhibition of protein biosynthesis, nucleic acid biosynthesis, protease activity and cell division.
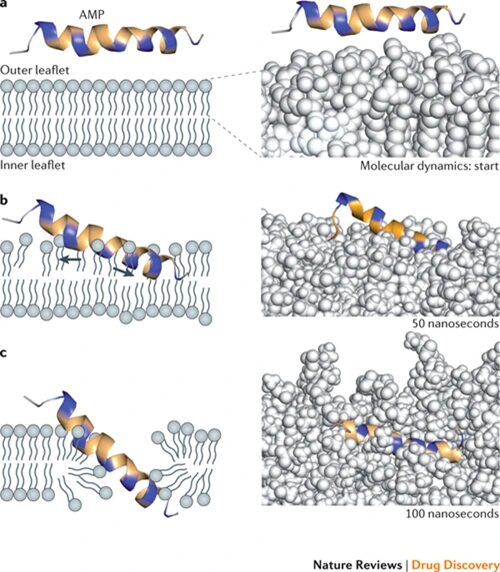
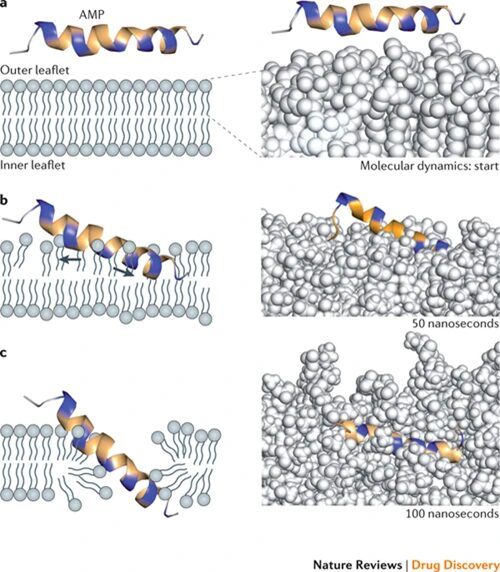
Fig. 14. Potential mechanism for membrane disruption and/or translocation. Positively charged amino acid residues indicated in blue, others in yellow. Right panels show computer simulated molecular dynamics: spinigerin in explicit water with a POPE (1-palmitoyl-2-oleoyl-phosphaethanolamine) membrane bilayer model. A) Initial membrane attachment. B) Stretching and thinning of the outer phospholipid layer. C) Insertion into the membrane (Fjell et al., 2012).
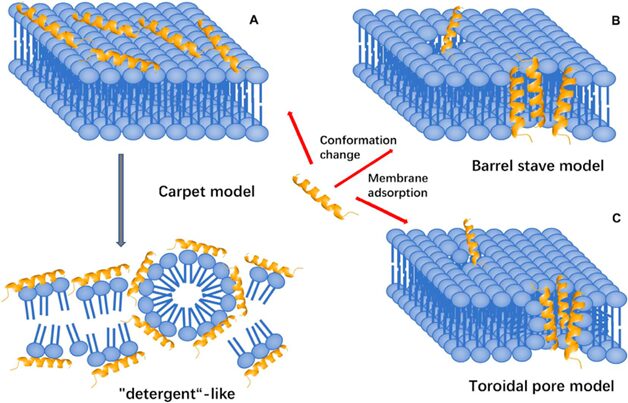
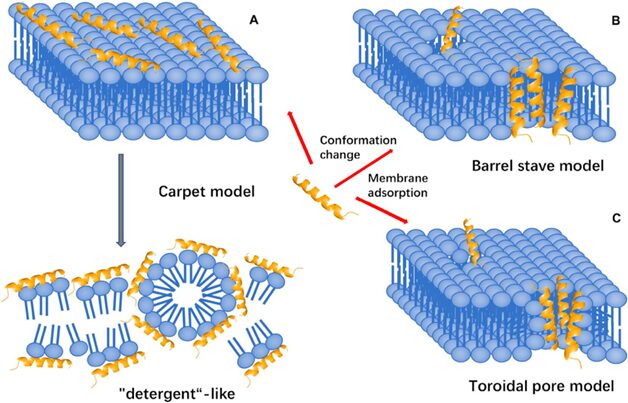
Fig. 15. Three membrane-targeting models (Huan et al., 2020).
Membrane targeting mechanisms
In the carpet model, the positively charged AMP initially interacts with the negatively charged lipid head groups of the outer surface of the cytoplasmic membrane. A high peptide concentration leads to increased membrane curvature and facilitates translocation across the membrane. The peptide is then inserted, in an approximately parallel orientation to the bilayer, into the outer layer of the membrane, leading to the displacement of lipids (Huan et al., 2020). In the barrel-stave model, AMPs aggregate, penetrate the membrane, and form channels resulting in cytoplasmic outflow. Lastly, in the toroidal pore model, AMPs penetrate and embed in the membrane, then bend to form a ring hole with a diameter of 1-2 nm. Several factors determine the interactions of an AMP with the membrane, including size, residue composition, overall charge, secondary structure, hydrophobicity, and amphiphilic character. Electrostatic attraction due to opposing charges or hydrophobic interactions alone is most likely insufficient for AMP activity (Fjell et al., 2012). Studies on the cell-penetrating peptide penetratin, a 16-residue cationic peptide which has been patented as a carrier for drug delivery into cells, have indicated the importance of AMPs’ molecular surface properties, as the mutation of a single amino acid residue abolished the membrane-transfer properties of penetratin.
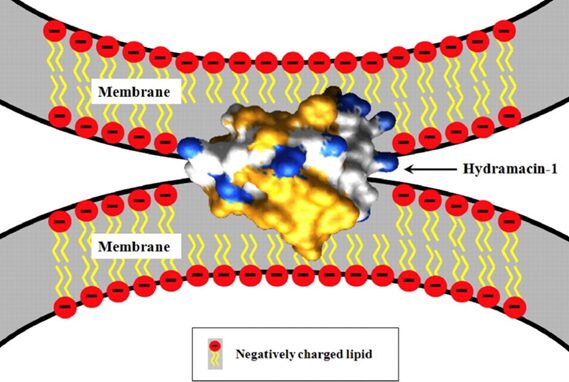
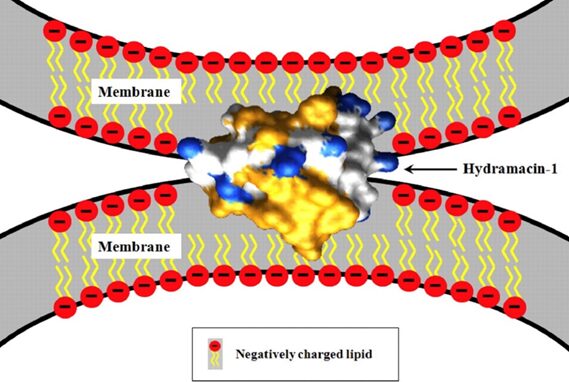
Fig. 16. Potential mechanism for bacterial aggregation by hydramacin-1, an AMP found in the epithelial cells of Hydrae. The AMP aggregates bacteria before destroying them through a yet unknown mechanism. Hydrophobic amino acid residues of the hydramacin-1 are indicated in yellow, while cationic surface areas are indicated in blue. Negative charges on the membrane surfaces of the two bacterial cells are believed to be compensated by the hydramacin-1’s positively charged belt running between its two hydrophobic ends (Jung et al., 2009).
Membrane composition preferences
Whether membrane binding represents the sole basis of AMP selectivity is unknown. However, there has been considerable discussion about how AMPs exhibit strong preferences for specific membrane compositions: contain comparatively large fractions of anionic lipids which attract cationic AMPs, maintain a high electrical potential gradient, and lack cholesterol (Fjell et al., 2012). These characteristics correspond to bacterial membranes, containing up to 20 mole percent of anionic lipids, maintaining a high electrical transmembrane potential of approximately -120 mV, and relatively low cholesterol compared to membranes of plant cells and animal cells.
AMPs in Hydrae
Currently, three families of AMPs have been identified in Hydra: the hydramacin, periculin, and arminin (Bosch, 2014). The arminin peptide family is an excellent choice for studying the role of AMPs in shaping the host-specific microbiota of Hydrae, due to its species-specific variability and elevated constitutive expression (constitutive expression refers to genes that are transcribed in an ongoing manner, with rate only influenced by metabolism). As previously mentioned, bacteria in Hydrae are specific for different host species. Closely related Hydra species are associated with similar microbial communities, and a high level of congruency can be observed between the evolution of the host species and the evolution of its associated microbiome (Bosch, 2014).
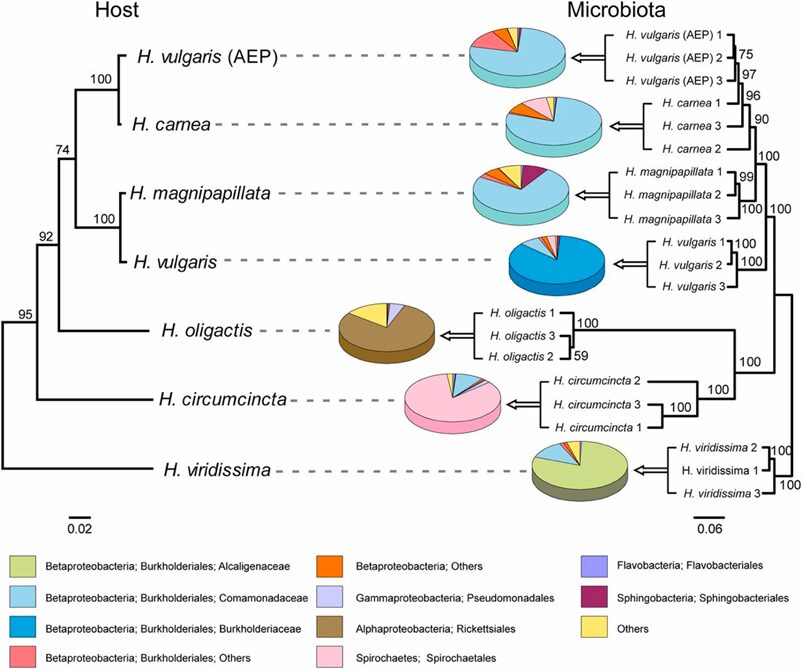
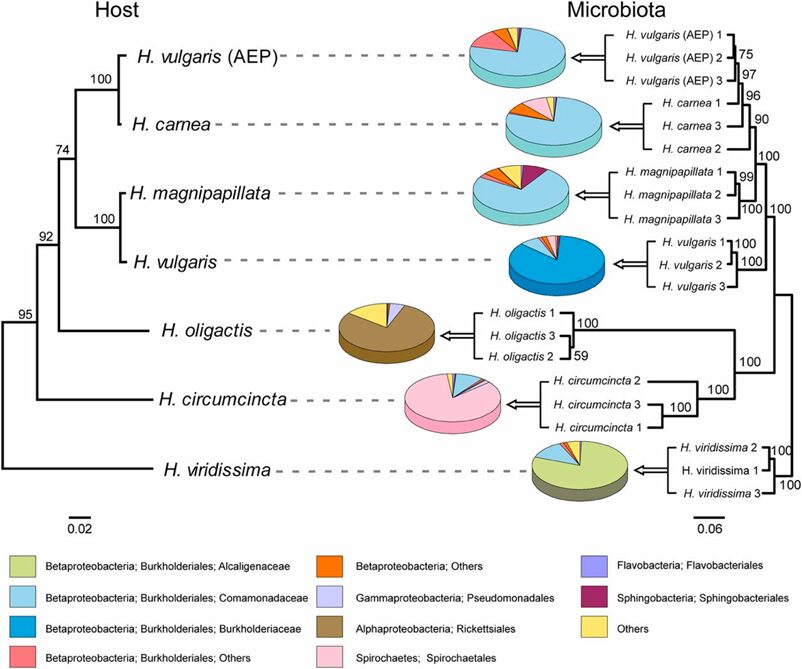
Fig. 17. Comparison of the phylogenetic tree of Hydra species and the cluster tree of the corresponding bacterial communities. Strong host-specificity of Hydra-associated bacterial communities, which reflects the phylogenetic relationship of their hosts (Franzenburg et al., 2013).
Species-specific microbiota
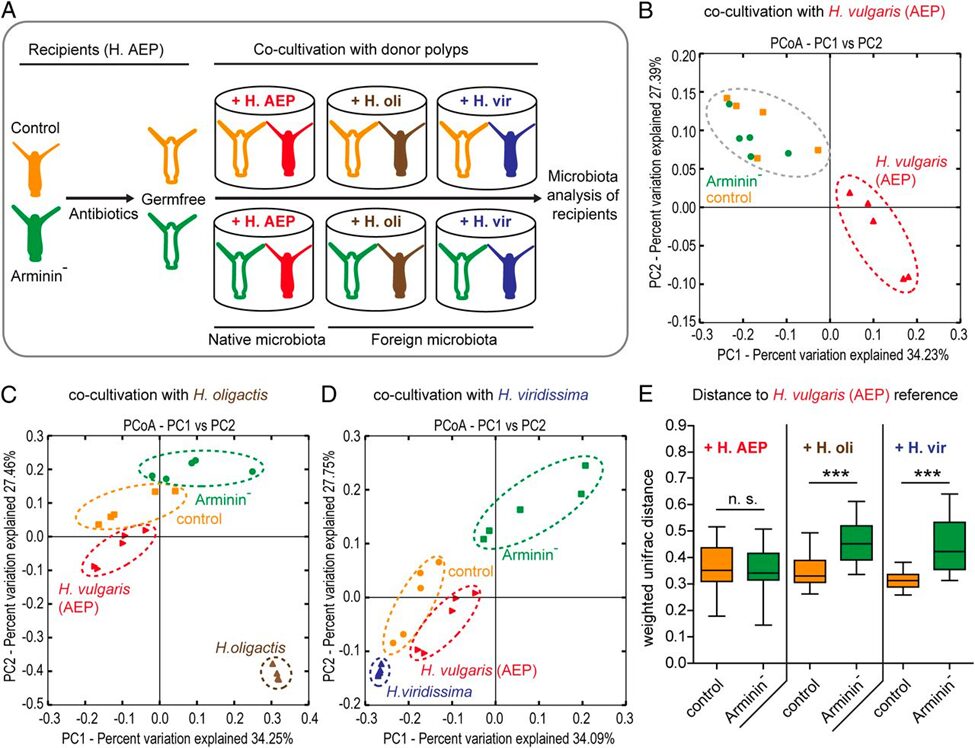
To confirm whether species-specific AMPs shape species-specific bacterial communities, a study examined the effect of arminin deficiency in Hydrae (Franzenburg et al., 2013). Arminin-deficient polyps and control polyps (both H. vulgaris) were generated by antibiotic treatment, and subsequently inoculated (introduced) to native bacterial communities (belonging to the same host species, H. vulgaris) as well as foreign bacterial communities (belonging to two closely related host species, H. oligactis and H. viridissima) by cocultivation with either of the three species. Both control and arminin-deficient polyps show no differences in bacterial recolonization when inoculated with native microbiota. However, when inoculated with foreign microbiota, control polyps selected for bacterial communities resembling their native microbiota, while this host-driven selection was significantly less pronounced in arminin-deficient polyps. Thus, it is confirmed that Hydra AMPs shape species-specific microbiota.
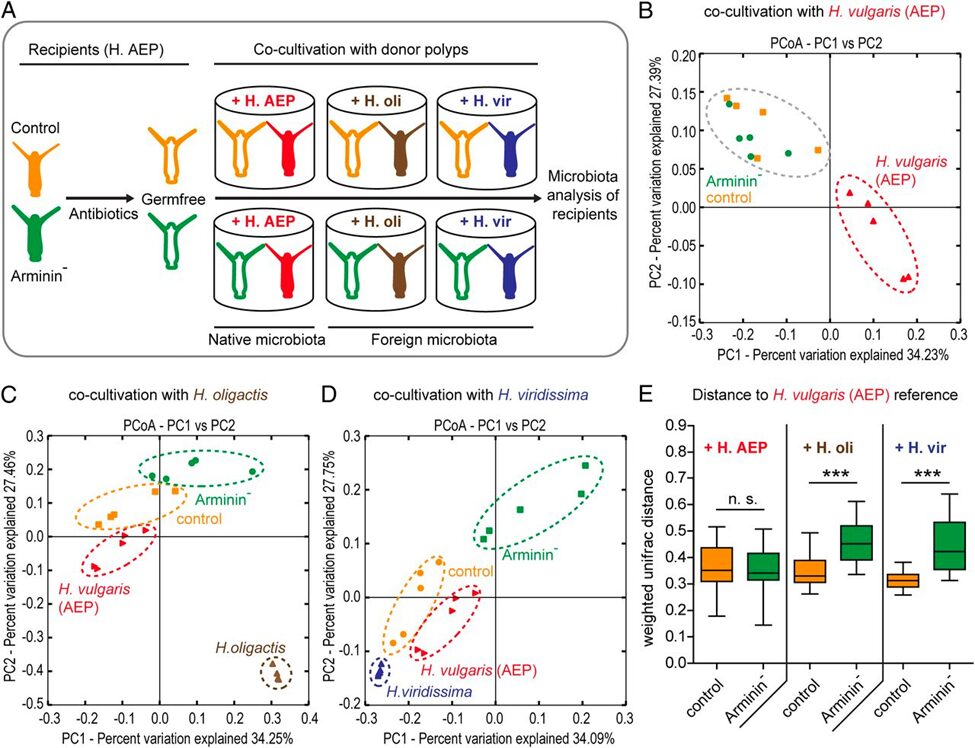
Fig. 18. A) Schematic representation of the experimental design. B) Cocultivation with the same Hydra species (native bacterial communities) results in similar bacterial recolonization for both the arminin-deficient and control polyps. C) and D) Cocultivation with different Hydra species results in separate clusters for the bacterial recolonization of arminin-deficient and control polyps. E) Control polyps have bacterial recolonization closer to H. oligactis and H. viridissima (different species) than arminin-deficient polyps, indicating a loss of selective preferences when arminin expression is disturbed (Franzenburg et al., 2013).
(3M, 2018; Loomis, 1959; Rachamim & Sher, 2012; Rangaraju et al., 2010; Rodrigues et al., 2016)
Fig. 19. Female polyp with developing embryo (right) and male poly with testes (left) (Stefan Siebert/Juliano Lab).
Mother to child AMPs transfer
An interesting note can be made on how Hydra transfers AMPs from mother to child, much like humans. Hydra mostly reproduce asexually by budding and reproduce sexually when conditions are harsh, and temperatures are low. In Hydra embryos, microbial colonization is controlled by maternally encoded AMPs. After mid-blastula transition, zygotically expressed AMPs take control of the microbiome, and a stable microbiome is established within 3 to 4 weeks which replaces the maternally produced AMPs (Bosch & Zasloff, 2021).
Since mutualistic associations between animals and beneficial microbes can increase host fitness, selective pressures act on host-mechanisms, which involves AMPs, to ensure advantageous bacterial colonization (Franzenburg et al., 2013). Furthermore, transmitted bacteria are selected for being beneficial to the host, given that they help increase the host’s survival rate. The interdependencies between the host and its associated microbes suggest that we should consider the metaorganism as a unit of natural selection. On the other hand, we have seen that in Hydrae, antimicrobial peptide profiles and the microbiota they help create are extremely diverse, even among closely related species. How and why could that be? Numerous studies have shown that the rapid evolution of AMPs observed in all types of organisms is encouraged by positive selection, where beneficial genetic variations sweep a population (Tennessen, 2005). It is possible that some hosts are in a constant co-evolutionary arms race with pathogens, and resistance is easy to evolve and happens frequently. Alternatively, AMP evolution might primarily occur when hosts enter new ecological niches, necessitating adaptation to entirely novel pathogen species they have not previously encountered. Given that Hydra species inhabit different habitats, it has been speculated that differences in their microbiota, shaped by diverse AMPs, contribute to diversification of their host species (Franzenburg et al., 2013).
Conclusion
Nature has come up with some of the most interesting design solutions in the millennia that it has been around. There is such a wealth of knowledge that can be obtained by observing and analyzing Hydra as we have done in our paper. We can learn from this tiny organism that will likely never interact with us in our environment by considering its solutions to the problems it faces. For example, the attachment of Hydrae to a surface addresses the problem of having very little control of its movement in a body of water with currents. Interestingly, attachment and detachment are not achieved by similar methods. The muscular action that detaches Hydrae from surfaces would not be useful for the attachment; therefore, Hydrae have evolved to secrete an adhesive material in order to attach itself. Additionally, one of Hydrae’s problems is that it may be small compared to its prey. Therefore, it evolved the ability to make its prey go limp after an attack. This allows for the Hydra to manipulate a larger creature and successfully eat it. The mesoglea plays a crucial role in maintaining tissue integrity and regeneration of Hydrae. Its capacity to form and reform rapidly during the regenerative process underscores its flexibility and adaptability to meet the organism’s needs. The organism’s unique chemical properties enable it to regenerate from dissociated cells efficiently. Since the organism lives in aquatic environments where physical damage and predation are common, regeneration increases their chance of survival and extends their lifespan. Due to Hydrae’s fragility, the organism regenerates instead of developing a form of physical protection like a shell. This highlights nature’s evolutionary design, where a simple yet versatile structure like the mesoglea is fine-tuned to play a central role in Hydrae’s vital regenerative abilities. Similarly, the innate immune system of Hydrae is also fine-tuned to efficiently protect the organism. Indeed, the renewable glycocalyx buffer at the ectodermal epithelium and the production of AMP induced by TLR-like receptors all work in junction to protect the animal from both sides in a way that takes advantage of its unique morphology and needs. Finally, the mechanisms of action of antimicrobial peptides, as well as their role in helping shape the most beneficial species-specific microbiome for the Hydra host is discussed. Chemical properties such as localized charges, as well as the structures and functions of AMP molecules, allow them to destroy bacterial cell membranes or aggregate bacteria for their subsequent elimination, as can be seen by the belted hydramacin-1. Transgenic arminin deficient polyps show the determining effect of specific AMP profile on bacterial composition, a process believed to have allowed diverse Hydra species to thrive in various aquatic environments.
References
3M. (2018). Technical Data Sheet 3M Scotch Transparent Film Tape. In.
Barzansky, B., & Lenhoff, H. M. (1974). On the chemical composition and developmental role of the mesoglea of Hydra. American Zoologist, 14(2), 575-581.
Barzansky, B. M. (1972). The mesolamella of hydra: on its composition, synthesis and developmental roles. University of California, Irvine.
Bosch, T. C. G. (2014). Rethinking the role of immunity: lessons from Hydra. Trends in Immunology, 35(10), 495-502. https://doi.org/https://doi.org/10.1016/j.it.2014.07.008
Bosch, T. C. G., Augustin, R., Anton-Erxleben, F., Fraune, S., Hemmrich, G., Zill, H., Rosenstiel, P., Jacobs, G., Schreiber, S., Leippe, M., Stanisak, M., Grötzinger, J., Jung, S., Podschun, R., Bartels, J., Harder, J., & Schröder, J.-M. (2009). Uncovering the evolutionary history of innate immunity: The simple metazoan Hydra uses epithelial cells for host defence. Developmental & Comparative Immunology, 33(4), 559-569. https://doi.org/https://doi.org/10.1016/j.dci.2008.10.004
Bosch, T. C. G., & Zasloff, M. (2021). Antimicrobial Peptides—or How Our Ancestors Learned to Control the Microbiome. mBio, 12(5), 10.1128/mbio.01847-01821. https://doi.org/doi:10.1128/mbio.01847-21
Epp, L., Smid, I., & Tardent, P. (1986). Synthesis of the mesoglea by ectoderm and endoderm in reassembled hydra. Journal of morphology, 189(3), 271-279.
Fjell, C. D., Hiss, J. A., Hancock, R. E. W., & Schneider, G. (2012). Designing antimicrobial peptides: form follows function. Nature Reviews Drug Discovery, 11(1), 37-51. https://doi.org/10.1038/nrd3591
Franzenburg, S., Walter, J., Künzel, S., Wang, J., Baines, J. F., Bosch, T. C. G., & Fraune, S. (2013). Distinct antimicrobial peptide expression determines host species-specific bacterial associations. Proceedings of the National Academy of Sciences, 110(39), E3730-E3738. https://doi.org/10.1073/pnas.1304960110
Fraune, S., & Bosch, T. C. (2010). Why bacteria matter in animal development and evolution. Bioessays, 32(7), 571-580. https://doi.org/10.1002/bies.200900192
Huan, Y., Kong, Q., Mou, H., & Yi, H. (2020). Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields [Review]. Frontiers in Microbiology, 11. https://doi.org/10.3389/fmicb.2020.582779
Jung, S., Dingley, A. J., Augustin, R., Anton-Erxleben, F., Stanisak, M., Gelhaus, C., Gutsmann, T., Hammer, M. U., Podschun, R., Bonvin, A. M., Leippe, M., Bosch, T. C., & Grötzinger, J. (2009). Hydramacin-1, structure and antibacterial activity of a protein from the basal metazoan Hydra. J Biol Chem, 284(3), 1896-1905. https://doi.org/10.1074/jbc.M804713200
Loomis, W. (1959). The sex gas of hydra. Scientific American, 200(4), 145-159.
Rachamim, T., & Sher, D. (2012). What Hydra can teach us about chemical ecology–how a simple, soft organism survives in a hostile aqueous environment. International Journal of Developmental Biology, 56(6-7-8), 605-611.
Rangaraju, S., Khoo, K., Feng, Z., Crossley, G., Nugent, D., & Khaytin, I. (2010). Chi v, Pham C, Calabresi P, Pennington Mw. In: Norton RS, Chandy KG.
Rau, I., & Kajzar, F. (2015). Biopolymers for application in photonics. Apple Academic Press Cambridge, MA, USA.
Rodrigues, M., Leclère, P., Flammang, P., Hess, M. W., Salvenmoser, W., Hobmayer, B., & Ladurner, P. (2016). The cellular basis of bioadhesion of the freshwater polyp Hydra. BMC Zoology, 1, 1-15.
Sarras Jr, M. P., Madden, M. E., Zhang, X., Gunwar, S., Huff, J. K., & Hudson, B. G. (1991). Extracellular matrix (mesoglea) of Hydra vulgaris: I. Isolation and characterization. Developmental biology, 148(2), 481-494.
Sarras Jr, M. P., Zhang, X., Huff, J. K., Accavitti, M. A., John, P. S., & Abrahamson, D. R. (1993). Extracellular matrix (mesoglea) of Hydra vulgaris: III. Formation and function during morphogenesis of hydra cell aggregates. Developmental biology, 157(2), 383-398.
Sato, M., & Sawada, Y. (1989). Regulation in the numbers of tentacles of aggregated hydra cells. Developmental biology, 133(1), 119-127.
Schröder, K., & Bosch, T. C. G. (2016). The Origin of Mucosal Immunity: Lessons from the Holobiont <i>Hydra</i>. mBio, 7(6), e01184-01116. https://doi.org/10.1128/mbio.01184-16
Technau, U., & Holstein, T. W. (1992). Cell sorting during the regeneration of Hydra from reaggregated cells. Developmental biology, 151(1), 117-127.
Tennessen, J. A. (2005). Molecular evolution of animal antimicrobial peptides: widespread moderate positive selection. Journal of Evolutionary Biology, 18(6), 1387-1394. https://doi.org/10.1111/j.1420-9101.2005.00925.x